В крахмал входит в состав: Крахмал состав — Справочник химика 21
Крахмал состав — Справочник химика 21
Минеральными удобрениями называют соли, содержащие элементы, необходимые для питания растений и вносимые в почву для получения высоких и устойчивых урожаев. В состав растений входят около 60 химических элементов. Для образования ткани растения, его роста и развития требуются в первую очередь углерод, кислород и водород, образующие основную часть растительной массы, далее азот, фосфор, калий, магний, сера, кальций и железо. Источниками веществ, необходимых для питания растений, служат воздух и почва. Из воздуха растения извлекают основную массу углерода в виде диоксида углерода, усваиваемого путем фотосинтеза, а из почвы — воду и минеральные вещества. Некоторое количество диоксида углерода воспринимается корневой системой растений из почвы. Среди минеральных веществ особенно важны для жизнедеятельности растений азот, фосфор и калий. Эти элементы способствуют обмену веществ в растительных клетках, росту растений и особенно плодов, повышают содержание ценных веществ (крахмала в картофеле, сахара в све-кле, фруктах и ягодах, белка в зерне), повышают морозостойкость и засухоустойчивость растений, а также их стойкость к заболеваниям.
Альдогексозы. о-Глюкоза (декстроза, виноградный сахар) — одно из наиболее распространенных органических соединений. Она содержится, например, во фруктах, растительных соках, лимфе, крови и т. д. Заметное количество о-глюкозы в моче наблюдается только при некоторых заболеваниях (диабете, или сахарной болезни). о-Глюкоза входит в состав многих олигосахаридов, полисахаридов и гликозидов.
 В промышленности получается гидролизом крахмала и служит сырьем для получения с помощью микробиальных процессов неко- [c.210]
В промышленности получается гидролизом крахмала и служит сырьем для получения с помощью микробиальных процессов неко- [c.210] Конденсация моносахаридов не ограничивается образованием дисахаридов. В живых организмах молекулы глюкозы могут конденсироваться тысячами, образуя гигантские молекулы. Входящие в их состав остатки глюкозы могут быть вытянуты в одну линию или же образовывать разветвленные цепи разной длины. Глюкоза входит в состав таких молекул, но только не в виде полных молекул, а в виде остатков, при соединении от каждых двух молекул глюкозы отщепляется по молекуле воды. Термин остаток применяют и к другим молекулам, соединяющимся путем конденсации в гигантские молекулы (их иногда называют макромолекулами). Такие гигантские молекулы имеет, например, крахмал. Он относится к полисахаридам ( много сахаров ). Конденсируясь с образованием крахмала, молекулы глюкозы теряют прежние свойства крахмал не растворяется в воде и несладок, он совершенно безвкусен.  145]
145]
Гидролизу подвергаются разные вещества соли, галогенан-пгдриды, карбиды, углеводы, белки, жиры и т. д. Разрушение горных пород обусловлено в значительной мере гидролизом составляющих их минералов — силикатов. В живых организмах происходит гидролиз белков, полисахаридов и других органических веществ. Состав и функция крови обусловлены гидролизом солей, растворенных в плазме. Осахаривание крахмала, гидролиз древесины, получение мыла и многие другие важные производства основаны иа гидролизе. [c.219]
Органические и неорганические высокомолекулярные соединения. Органические высокомолекулярные соединения являются основой живой природы. Важнейшие соединения, входящие в состав растений, — полисахариды, лигнин, белки, пектиновые вещества — высокомолекулярны. Ценные механические свойства древесины, хлопка, льна обусловлены значительным содержанием в них высокомолекулярного полисахарида— целлюлозы. Главной составной частью картофеля, пшеницы, ржи, овса, риса, кукурузы, ячменя является другой высокомолекулярный полисахарид — крахмал. Торф, бурый уголь, каменные угли представляют собой продукты геологического превращения растительных тканей, главным образом целлюлозы и лигнина, и также должны быть отнесены к высокомолекулярным соединениям. [c.11]
Торф, бурый уголь, каменные угли представляют собой продукты геологического превращения растительных тканей, главным образом целлюлозы и лигнина, и также должны быть отнесены к высокомолекулярным соединениям. [c.11]
Углеводы в форме крахмала являются важнейшими источниками энергии в пище. Для получения этой энергии мы либо употребляем в пищу зерна, в которых накапливается крахмал, либо скармливаем эти зерна животным, которые синтезируют мясные белки, а затем съедаем их. В любом случае потребляемая нами энергия в конце концов поставляется крахмалом, полимерным продуктом фотосинтеза. Целлюлоза входит в состав хлопка и льна, а также искусственных продуктов — ацетата целлюлозы и вискозного волокна. Дерево, из которого сделана наша мебель, также содержит целлюлозу. Бумага этой книги получена в процессе обработки целлюлозы. Даже деньги давно перестали делать из благородных металлов, заменив их целлюлозой.

В состав крахмала входят два полисахарида амилоза (20— 30%) и амилопектин (70—80%). Эти полисахариды построены из остатков а- )-глюкозы [c.100]
По своему строению водоросли могут быть одноклеточными, многоклеточными и колониальными формами. Некоторые из них имеют клетку без плотной оболочки и лишь с уплотненным внешним слоем протоплазмы, вследствие чего обладают способностью изменять свою форму. Другие же характеризуются плотной оболочкой, большей частью состоящей из целлюлозы. Часто в состав оболочки входят пектиновые вещества. У некоторых групп оболочка сильно пропитана известью или кремнеземом. Одни клетки содержат одно или несколько ядер, другие типичного ядра не имеют, лишь в протопласте заметна окрашенная периферическая часть и неокрашенное центральное тело. У некоторых водорослей красящие вещества находятся в особых плазменных телах различной формы, которые называются хроматофорами. Большей частью в хроматофоры бывают включены плотные тельца — пиреноиды, богатые белковыми веществами. Вокруг пиреноидов отлагается крахмал, являющийся одним из продуктов ассимиляции. Запасными питательными веществами служат масла, жиры, лейкозин, маннит и глюкоза.
Большей частью в хроматофоры бывают включены плотные тельца — пиреноиды, богатые белковыми веществами. Вокруг пиреноидов отлагается крахмал, являющийся одним из продуктов ассимиляции. Запасными питательными веществами служат масла, жиры, лейкозин, маннит и глюкоза.
Для придания огнестойкости ткани рекомендуется такой состав сульфата аммония 8 мае. ч., борной кислоты 3 мае. ч., буры 2 мае. ч., крахмала 2 мае. ч., воды 85 мае. ч. Сначала готовят клейстер из крахмала и воды, затем в свежеприготовленном клейстере растворяют остальные составные части и погружают в него ткань. Одновременно 125 г хозяйственного мыла растворяют в 12 л горячей (90°С) воды и 165 г квасцов растворяют в 12 л тоже нагретой до 90°С воды. Ткань, отжатую от первого (крахмального) раствора, несколько раз погружают в мыльную воду, затем в раствор квасцов и сушат на воздухе. [c.76]
Считают (см. доп. [1], стр. 434), что комплекс с иодом образует входящая в соста крахмала амилоза.
Глюкоза (виноградный сахар) СвН аОв (стр. 223). Одна из наиболее часто встречающихся в природе альдогексоз. ( Содержится в соке винограда и других плодов, а также (вместе с фруктозой) в меде. Входит в состав крови и других биологических жидкостей животных организмов. Является составной частью многих полисахаридов, из которых и может быть получена при гидролизе. В технике О-глюкозу получают гидролизом крахмала в присутствии минеральных кислот (стр. 262). Чистая О-глюкоза получается из так называемого инвертного сахара (стр. 258) — смеси О-глюкозы и О-фруктозы, образующейся при гидролизе тростникового сахара разделение этих моносахаридов основано на их различной растворимости в спирте. [c.247]
Крахмал — широко распространенный в природе полисахарид, макромолекула которого состоит из звеньев, представляюш,их собой остатки О-глюкозы.
Организмы животных содержат сравнительно немного углеводов.. Напротив, в растениях они образуют основную массу тканей. Последние состоят главным образом из клетчатки, имеющей тот же состав, что и крахмал, но еще больший молекулярный вес. [c.541]
При действии хлористого сульфурила на крахмал в разбавленном растворе щелочи [311] получается смесь соединений, содержащих небольшие количества серы. Молекулярные веса этих веществ свидетельствуют о глубокой деградации молекулы крахмала. Обработка хлорсульфоновой кислотой в пиридине ведет к образованию кислого сульфата крахмала [312], из которого выделена калиевая соль, имеющая состав СвНд05(80зК)2 2,бНзО. [c.56]
Сырье пищевой промышленности имеет растительное и животное происхождение и содержит разнообразные органические вещества. Ценными питательными продуктами являются различные растительные и животные жиры, углеводы — разнообразные сахара, крахмал, патока и др.
Углеводы — вещества состава С Н2 0 , имеющие первостепенное биохимическое значение, широко распространенные в живой природе. К ним относятся различные сахаристые вещества, крахмал, целлюлоза (клетчатка). Название углеводы сохранилось за ними с тех времен, когда строение этих соединений еще не было изучено, но установлен был их состав, отвечающий общей формуле С (Н20) . По этой формуле углеводы рассматривали как гидраты углерода — соединения углерода с водой — углеводы.
Имея тот же химический состав, что и клетчатка, крахмал по внешнему виду сильно отличается от нее. Вместо волокон он образует белые зерна диаметром в несколько десятков микрон.
 Крахмал нерастворим в холодной воде, в горячей же набухает и постепенно растворяется. Образующиеся вязкие растворы при понижении температуры превращаются в студневидную массу — клейстер. [c.308]
Крахмал нерастворим в холодной воде, в горячей же набухает и постепенно растворяется. Образующиеся вязкие растворы при понижении температуры превращаются в студневидную массу — клейстер. [c.308]Большую часть вырабатываемого крахмала составляет картофельный крахмал, так как картофель—широко распространенная культура, дающая высокие урожаи. При хорошем уходе с])едний урожай картофеля составляет 200 ц га, что в пересчете на yxoir крахмал состав.ляот около 40 цгга картофельного крахмала. Необходимо отметить, что развивающееся социалистическое сельское хозяйство добилось неслыханных урожаев картофеля—свы-Hie 1000 ц га. [c.123]
Для увеличения производительности сварки и улучшения качества сва рного шва рекомендуют присадочные чугунные прутки предварительно обмазывать флюсом, замешенным на жидком стекле или водном растворе крахмала. Состав флюса 57% технической буры (ГОСТ 8429—69) 29% плавикового шпата (ГОСТ 4421—48) 7% ферросилиция ФС75 (ГОСТ 1415—70). Кроме того, в состав флюса вводят 2%-ный раствор пищевого крахмала в количестве 100% к массе смеси сухих компонентов флюса. При составлении смеси указанных веществ нх просеивают через сито 600—900 0твс рстий на 1 см . [c.54]
Кроме того, в состав флюса вводят 2%-ный раствор пищевого крахмала в количестве 100% к массе смеси сухих компонентов флюса. При составлении смеси указанных веществ нх просеивают через сито 600—900 0твс рстий на 1 см . [c.54]
Вёлер, Кольбе и Бертло синтезировали относительно простые органические соединения, тогда как для живой природы характерны значительно более сложные соединения типа крахмала, жиров и белков. Изучать такие соединения гораздо труднее непросто даже установить их точный элементный состав. В целом изучение органических веществ обещало разгадку многих проблем, но подступиться к этим веществам химику прошлого века было совсем непросто. [c.71]
Полисахариды. Эти углеводы во многом отличаются от моно- и дисахаридов — не имеют сладкого вкуса, в большинстве нерастворимы в воде, они представляют собой сложные высокомолекулярные соединения, которые под каталитическим влиянием кислот или ферментов подвергаются гидролизу с образованием более простых полисахаридов, затем дисахаридов и, в конечном итоге, множества (сотен и тысяч) молекул моносахаридов. Важнейшие представители полисахаридов — крахмал и целлюлоза (клетчатка). Их молекулы построены из звеньев -СбНюОб-, являющихся остатками шестичленных циклических форм молекул глюкозы, потерявших молекулу воды поэтому состав и крахмала, и целлюлозы выражается общей формулой (СеНюОа) . Различие же в свойствах этих полисахаридов обусловлено пространственной изомерией образующих их моно-сахаридных молекул крахмал построен из звеньев а-, а целлюлоза — /3-формы глюкозы. [c.582]
Важнейшие представители полисахаридов — крахмал и целлюлоза (клетчатка). Их молекулы построены из звеньев -СбНюОб-, являющихся остатками шестичленных циклических форм молекул глюкозы, потерявших молекулу воды поэтому состав и крахмала, и целлюлозы выражается общей формулой (СеНюОа) . Различие же в свойствах этих полисахаридов обусловлено пространственной изомерией образующих их моно-сахаридных молекул крахмал построен из звеньев а-, а целлюлоза — /3-формы глюкозы. [c.582]
Полисахариды. Эти углеводы во многом отличаются от MOHO- и дисахаридов — не имеют сладкого вкуса, в большинстве нерастворимы в воде они представляют собой сложные высоко-мо.г1екулярные соединения, которые под каталитическим влиянием кислот или ферментов подвергаются гидролизу с образованием более простых полисахаридов, затем дисахаридов и, в конечном итоге, множества (сотен и тысяч) молекул моносахаридов. Важнейшие представители полисахаридов — крахмал и целлюлоза (клетчатка). Их молекулы построены из звеньев — eHioOj—, являющихся остатками шестичленных циклических форм молекул глюкозы, потерявших молекулу воды поэтому состав и крахмала, [c. 493]
493]
С,Н120в — самый распространенный моносахарид (углевод). Встречается в свободном состоянии особенно много ее в еиноградном соке, откуда другое название Г.— виноградный сахар. Г. входит в состав молекул крахмала, целлюлозы, декстрина, гликогена, мальтозы, сахарозы и многих других ди- и полисахаридов, из которых Г. получают как конечный продукт гидролиза. В печени человека из Г. синтезируется гликоген, в промышленности Г. получают гидролизом крахмала или клетчатки. При восстановлении Г. образуется шестиатомный спирт сорбит. Г. легко окисляется, дает реакцию серебряного зеркала. Г. широко применяется в медицине как вещество, легко усваивающееся организмом, при сердечных заболеваниях, шоковом состоянии, после операций. Г. [c.78]
Процесс образования угля в природе, называемый углефикацией или карбонизацией, разделяется на биохимическую (диагенезис) и геологическую (метаморфизм) стадии [63], На стадии диагенезиса углеводородные соединения растительных остатков (целлюлоза, лигнин, глюкоза, крахмал и др.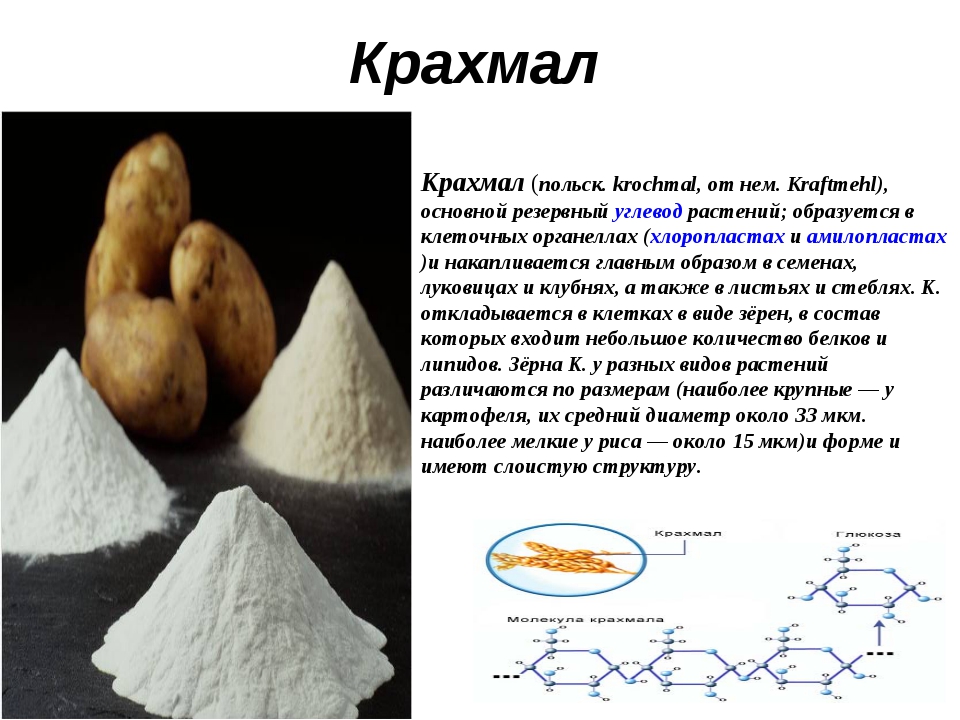 ) в результате реакций окисления кислородом воздуха и кислородом, содержащимся в проточных водах, а также под воздействием анаэробных бактерий превращались в гомогенизированное вещество — гумус. Б гумусе продолжалось взаимодействие входящих в его состав органических и привнесенных водой неорганических компонентов. Стадия метаморфизма проходила лосле образования над отложившейся органической массой достаточно мощных осадочных слоев неорганических веществ, т. е. на большой глубине и при высоких давлениях и температурах без доступа воздуха. В таких условиях органическое вещество уплотнялось и обезвоживалось, из него выделялся метан, что приводило к уменьшению содержания кислорода и водорода и росту содержания углерода. [c.64]
) в результате реакций окисления кислородом воздуха и кислородом, содержащимся в проточных водах, а также под воздействием анаэробных бактерий превращались в гомогенизированное вещество — гумус. Б гумусе продолжалось взаимодействие входящих в его состав органических и привнесенных водой неорганических компонентов. Стадия метаморфизма проходила лосле образования над отложившейся органической массой достаточно мощных осадочных слоев неорганических веществ, т. е. на большой глубине и при высоких давлениях и температурах без доступа воздуха. В таких условиях органическое вещество уплотнялось и обезвоживалось, из него выделялся метан, что приводило к уменьшению содержания кислорода и водорода и росту содержания углерода. [c.64]
Полисахариды (полимерные углеводы) представляЕот собой соединения, состоящие из многих сотен нли даже тысяч моносаха-ридных звеньев. Их состав отвечает общей формуле (СеНюОз) . Наиболее важными среди полисахаридов являются целлюлоза и крахмал. Оба эти вещества образуются в растениях из диоксида углерода и воды в результате фотосинтеза. Целлюлоза — основной строительный материал растений, крахмал служит запасным пищевым фондом растений и находится в основном в семенах (кукуруза, картофель, рис, пшеница и др.). Углеводы служат источником питания человека. В организме человека и животных они превращаются в жиры и белки. Целлюлоза в виде хлопка и вискозы применяется для изготовления одежды и бумаги. [c.307]
Оба эти вещества образуются в растениях из диоксида углерода и воды в результате фотосинтеза. Целлюлоза — основной строительный материал растений, крахмал служит запасным пищевым фондом растений и находится в основном в семенах (кукуруза, картофель, рис, пшеница и др.). Углеводы служат источником питания человека. В организме человека и животных они превращаются в жиры и белки. Целлюлоза в виде хлопка и вискозы применяется для изготовления одежды и бумаги. [c.307]
УГЛЕВОДЫ (глюциды, глициды)—важнейший класс органических соединений, распространенных в природе, состав которых соответствует общей формуле С (НзО) — По химическому строению У.— альдегидо- или кетоноспирты. Различают простые У.— моносахариды (сахара), например глюкоза, фруктоза, и сложные—полисахариды, которые делят на низкомолекулярные У.— дисахариды (сахароза, лактоза и др.) и высокомолекулярные, такие, например, как крахмал, клетчатка, гликоген. Характерным для У. является то, что моносахариды не гидролизуют, а молекулы полисахаридов при гидролизе расщепляются на две молекулы (дисахариды) или на большее число молекул (крахмал, клетчатка) моносахаридов. У. имеют огромное значение в обмене веществ организмов, являясь главным источником [c.255]
У. имеют огромное значение в обмене веществ организмов, являясь главным источником [c.255]
Состав и содержание твердой, жидкой и газообразной фаз в буровом растворе важно знать особенно при разбуривании обваливающихся горных пород. Регулируя состав фильтрата и pH раствора, можно эффективно бороться с обвалообразованиями. Изменение pH фильтрата в процессе бурения указывает на вероятность каких-либо отклонений величин других показателей раствора от установленных. При высоких значениях pH подавляются процессы ферментативного разложения содержащихся в растворах органически х реагентов типа крахмала [154, 169]. [c.39]
В промывочной жидкости реакции деструкции могут также протекать под действием химических агентов (химическая деструкция) и механических воздействий (механическая деструкция). Химическая деструкция протекает под действием полярных веществ воды, кислорода, щелочей, кислот и т. д. Наиболее распространении ми ее видами являются гидролиз и окислительная деструкция. При гидролизе бо1совых функциональных групп изменяется химическим состав полимера (например, гидролизованные полиакрил-онитрил и полиакриламид). При гидролизе же связей, входящих в состав основной цепи, происходит собственно деструкция с умс1и,шеннем средней молекулярной массы полимера, как, например, в кислой среде у крахмала — [c.36]
При гидролизе бо1совых функциональных групп изменяется химическим состав полимера (например, гидролизованные полиакрил-онитрил и полиакриламид). При гидролизе же связей, входящих в состав основной цепи, происходит собственно деструкция с умс1и,шеннем средней молекулярной массы полимера, как, например, в кислой среде у крахмала — [c.36]
В случае необходимости получения ванны с высокой плотностью можно применять медный купорос, карбонат меди, гидраты окиси меди, а в качестве стабилизаторов — КМЦ-600, карбофен, крахмал и др. Для глушения газового пласта в скв. 10 Чуст-Пап была предложена ванна, в состав которой входило 10% медного купороса, 15% глинопорошка, 10% буры, 1,2% карбофена и барит. Показатели ванны имели следующие значения = 1,64 — -г 1,66 г/смз, Бзо = 4 м Т = 40 45 с, pH = 7,0 — 7,5. [c.268]
Гексозы (СбН120б). D-глюкоза (виноградный сахар) — кристаллическое вещество, хорошо растворимое в воде, i n безводной глюкозы равна 146°С. Примерно в два раза уступает по сладости сахарозе. Встречается в растениях в свободном виде, а также входит в состав ди- и полисахаридов. В промышленности глюкозу получают из крахмала кипячением с разбавленной серной кислотой. [c.243]
Примерно в два раза уступает по сладости сахарозе. Встречается в растениях в свободном виде, а также входит в состав ди- и полисахаридов. В промышленности глюкозу получают из крахмала кипячением с разбавленной серной кислотой. [c.243]
Крахмал — самый распространенный в природе полисахарид, играющий роль резервного продукта многих растений. В технике крахмал, в основном, получают нз картофеля. В состав крахмала входят два полисахарида — ажилоза (20—30%) и амилопектин (70—80%). Эти полисахариды построены из остатков а-О-глюко-зы, связанных между собой а-(1,4 )-глюкозидными связями [c.247]
ЗУБНОЙ ПОРОШОК — гигиеническ(к средство для чистки зубов и полости рта состоит оно в основном из карбоната кальция (мела), иногда с примесью до 10% карбоната магния. Для вкуса и запаха добавляют мятное масло, иногда вместе с анисовым, гвоздичным, коричным и другими, а также с ментолом. Кроме этих составных частей, в состав зубной пасты входят водно-гли-цериновые растворы агар-агара, крахмала, мыла и других веществ, придаюгцих пластичность. 3. п. полирует зубную эмаль и нейтрализует кислоты, отравляющие ее. [c.101]
3. п. полирует зубную эмаль и нейтрализует кислоты, отравляющие ее. [c.101]
КРАХМАЛ ( 5Hi U5) — полисахарид, образуется в листьях растений в результате фотосинтеза и откладывается в корнях, клубнях, семенах в виде зерен, характерных для каждого вида растений. В состав К. входят амилоза и амилопек-тин. К- дает синее окрашивание с иодом, подвергается гидролизу, в результате чего образуется глюкоза [c.137]
Клетчатка, или целлюлоза, (СбНю05)п имеет такой же химический состав, как и крахмал. Она отличается от последнего структурой макромолекулы у крахмала разветвленная структура молекул, а молекулы клетчатки линейны. [c.360]
Крахмал — белый аморфный порошок, напоминающий муку (картофельная мука),-нераегворимый в холодной воде. В горячей воде набухает, образуя внешне однородный раствор крахмального клейстера. Состав крахмала выражается формулой (СвН, Ов) . [c.165]
Приблизительный состав ультрамарина отвечает формуле NagAleSieSgOgi. Ультрамарин применяют в качестве синьки для белья, для окраски стен, а также в производстве бумаги, сахара и крахмала для придания белоснежного цвета этим продуктам. [c.492]
Ультрамарин применяют в качестве синьки для белья, для окраски стен, а также в производстве бумаги, сахара и крахмала для придания белоснежного цвета этим продуктам. [c.492]
Молекулы углеводов наряду с несколькими группами ОН содержат обычно группу СНО. В связи с этим для углеводов одновременно характерны свойства и спиртов, и альдегидов. Само название этого класса веществ, к числу которых относятся, в частности, такие важные продукты питания, как обыкновенный сахар ( i2h320n) и крахмал [( eHioOs) ас], связано с тем, что их водород и кислород находятся обычно в таком же соотношении, как в воде (т. е. атомов водорода вдвое больше, чем кислорода). Поэтому состав громадного большинства углеводов может быть выражен в виде пС + тНаО (т. е. как бы в виде соединения угля с водой). [c.541]
Молекулы крахмала, гликогена и целлюлозы построены из пира-нозпых (с шестичленным кольцом) звеньев О-глюкозы последняя является конечным продуктом гидролиза всех этих полисахаридов, поэтому состав любого из них выражается общей формулой (СвНюОй) , где л — величина порядка сотен и тысяч. [c.259]
[c.259]
Строение молекул целлюлозы. Так же как молекулы крахмала, молекулы целлюлозы представляют собой цепи, построенные из остатков О-глюкозы в пиранозной форме, каждый из которых своим полуацетальным (1-м) углеродом через кислород соединен с 4-м углеродным атомом следующего глюкозного остатка. Основное различие в строении полисахаридов крахмала и целлюлозы заключается в том, что в состав последних глюкоза входит не в а-, а в Р-пиранозной форме. Таким образом, цепи целлюлозы построены по типу дисахарида целлобиозы (стр. 253) и представляют собой полигликозиды со множеством Р-1,4-гликозидных связей. Строение цепи молекулы целлюлозы наиболее наглядно выражается перспективной формулой [c.263]
Глюкоза была впервые получена в 1811 г. русским химиком К. С. Кирхгофом при гидролизе крахмала. В 60-х годах прошлого столетия глюкозе приписывали эмпирическую формулу СвН Ог. Известно было, что она легко теряет молекулу воды, приобретая состав gHiaOe. Тогда еще не знали, что при этом просто удаляется кристаллизационная вода. Из того факта, что глюкоза образует сложные эфиры с органическими кислотами, был сделан правильный вывод, что в глюкозе содержатся гидроксильные группы. [c.282]
Тогда еще не знали, что при этом просто удаляется кристаллизационная вода. Из того факта, что глюкоза образует сложные эфиры с органическими кислотами, был сделан правильный вывод, что в глюкозе содержатся гидроксильные группы. [c.282]
Глюкоза входит также в состав важнейших природных ди- и полисахаридов сахарозы, мальтозы, лактозы, клетчатки, крахмала. Довольно распространены в природе и некоторые глюкозиды, роль спиртового компонента (аглюкона) в которых могут играть такие соединения, как фенолы, циангидрины альдегидов и др. К глюко-зидам относятся, в частности, красящие вещества растений, обладающие сильным физиологическим действием сердечные глюкозиды, дубильные вещества. Примером может служить глюкозид амигда-лин .2oh3,0,iN. Он содержится в зернах горького миндаля и косточках других плодов. По своему строению он является глюкозидом дисахарида генциобиозы и циангидрина бензальдегида. При гидролизе кислотами амигдалин распадается на компоненты [c.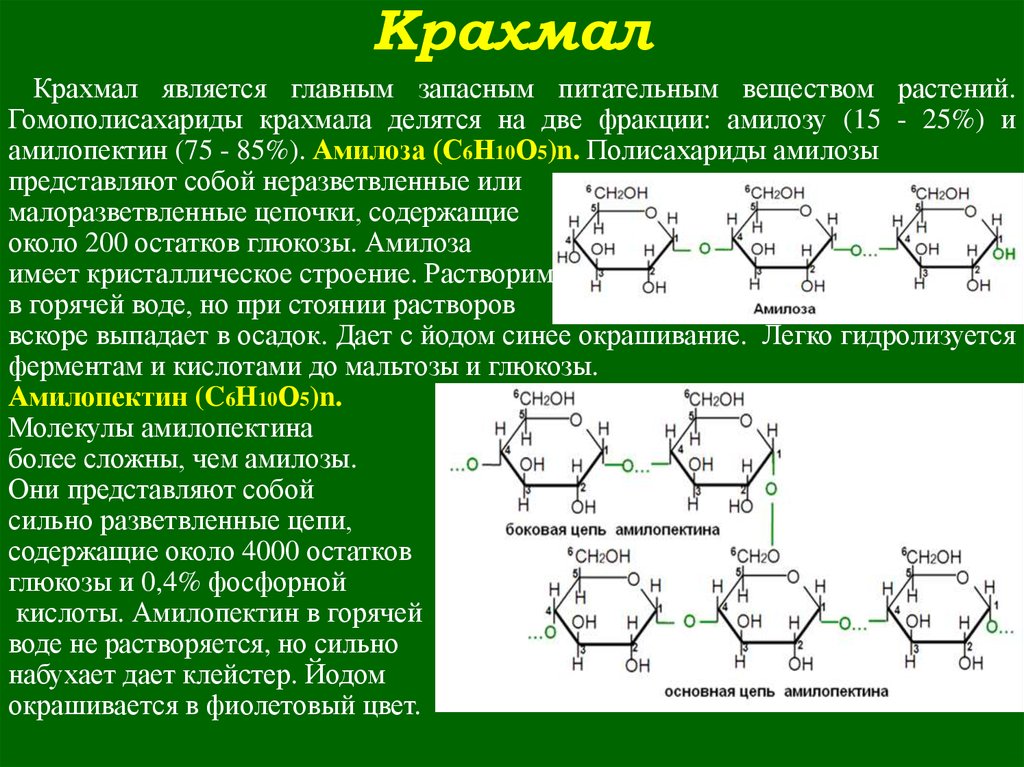 302]
302]
С участием третьей молекулы моносахарида может образовываться трисахарид С1аНзо01в, затем тетрасахарид СмИ гОг и т. д. По мере роста числа звеньев состав соединения будет соответствовать формуле (СвНюОа) . Эта и есть формула полисахаридов, из которых важнейшие — клетчатка и крахмал. [c.315]
Крахмал — урок. Химия, 8–9 класс.
Полисахариды — углеводы, в молекулах которых многократно повторяются остатки циклической формы глюкозы.
Состав всех полисахаридов выражается формулой C6h20O5n. Все полисахариды подвергаются гидролизу, продуктом которого является глюкоза:
C6h20O5n+nh3O→nC6h22O6.
К полисахаридам относятся крахмал, гликоген (животный крахмал) и целлюлоза (клетчатка).
Крахмал является запасным веществом растений и широко распространён в природе. Он в больших количествах содержится в зёрнах ржи, пшеницы, кукурузы, в клубнях картофеля.
Образуется крахмал в клетках растений из глюкозы:
nC6h22O6→C6h20O5n+nh3O.
Число повторяющихся структурных звеньев в макромолекуле крахмала может колебаться от \(100\) до \(6000\). Относительная молекулярная масса молекул тоже сильно различается.
Макромолекулы крахмала имеют линейное и разветвлённое строение.
Рис. \(1\). Молекулы крахмала
Крахмал — белое аморфное вещество, нерастворимое в холодной воде. В горячей воде он набухает и образует клейстер. Не имеет вкуса.
Рис. \(2\). Крахмал
Характерная реакция крахмала — реакция с иодом. При действии раствора иода на крахмал образуется соединение синего цвета. Эту реакцию используют для качественного определения крахмала.
Рис. \(3\). Действие иода на крахмал
Крахмал входит в состав продуктов питания и является основным источником углеводов в организме человека.
В организме крахмал подвергается гидролизу под действием пищеварительных ферментов. Процесс переваривания крахмала начинается уже в ротовой полости под действием ферментов слюны. Далее гидролиз крахмала продолжается в желудке и в кишечнике. Образовавшаяся глюкоза всасывается в кровь, а затем используется клетками для получения энергии.
Крахмал применяется для получения глюкозы. Используется для проклейки бумаги и картона, для пропитки тканей. Применяется он также при изготовлении таблеток в качестве наполнителя.
Источники:
Рис. 1. Молекулы крахмала https://image.shutterstock.com/image-vector/illustration-chemical-polysaccharide-form-which-600w-1738186163.jpg
Рис. 2. Крахмал
Рис. 3. Действие иода на крахмал
Крахмал — это… Что такое Крахмал?
Крахмал — Крахмал … Википедия
КРАХМАЛ — запасной углевод большинства растений; находится в них в виде крахмальных зёрен. Выделенный из растений крахмал безвкусный, белый, скрипящий между пальцами порошок, по внешнему виду напоминает муку (отсюда название крахмала, полученного из… … Краткая энциклопедия домашнего хозяйства
Выделенный из растений крахмал безвкусный, белый, скрипящий между пальцами порошок, по внешнему виду напоминает муку (отсюда название крахмала, полученного из… … Краткая энциклопедия домашнего хозяйства
крахмал — (от нем. Kraftmehl крепкая, сильная мука). Мучнистое, сильное клеющее вещество ряда растений. В кулинарном и кондитерском деле применяют чаще всего картофельный, пшеничный и кукурузный крахмал, причем последний наиболее нежный. Крахмал идет… … Кулинарный словарь
Крахмал — природный углевод, накапливаемый в клетках растений в виде крахмальных зерен и выделяемый из крахмалсодержащего сырья при его переработке;… Источник: Постановление Правительства РФ от 09.03.2010 N 132 Об обязательных требованиях в отношении… … Официальная терминология
КРАХМАЛ — Amylum. Крахмал полисахарид, получаемый из зерен пшеницы, кукурузы, риса, клубней картофеля. Свойства. Представляет собой белый мучнистый порошок без запаха и вкуса. Нерастворим в холодной воде, в горячей воде образует коллоидный раствор (клейст … Отечественные ветеринарные препараты
Нерастворим в холодной воде, в горячей воде образует коллоидный раствор (клейст … Отечественные ветеринарные препараты
КРАХМАЛ — КРАХМАЛ, (С6Н10О5)п, углевод, относящийся к коллоидным полиозам. Конечным продуктом при кислотном его гидролизе является й глюкоза, при ферментативном (амилаза) мальтоза; промежуточными продуктами гидролиза являются растворимый К и декстрины… … Большая медицинская энциклопедия
КРАХМАЛ — (польское krochmal, от немецкого Kraftmehl), запасной углевод растений; состоит из 2 полисахаридов амилозы и амилопектина, образованных остатками глюкозы. Накапливается в виде зерен, главным образом в клетках семян, луковиц, клубней, а также в… … Современная энциклопедия
КРАХМАЛ — (польск. krochmal от нем. Kraftmehl), запасной углевод растений; состоит из двух полисахаридов амилозы и амилопектина, образованных остатками глюкозы. Накапливается в виде зерен, главным образом в клетках семян, луковиц, клубней, а также в… … Большой Энциклопедический словарь
КРАХМАЛ — КРАХМАЛ, УГЛЕВОД, накапливающийся во многих растениях; он составляет около 70% питания человечества, поскольку содержится в таких продуктах, как картофель, пшеница, рис и другие злаки. Растения и животные преобразуют крахмал в ГЛЮКОЗУ, которая… … Научно-технический энциклопедический словарь
Растения и животные преобразуют крахмал в ГЛЮКОЗУ, которая… … Научно-технический энциклопедический словарь
КРАХМАЛ — КРАХМАЛ, крахмала, мн. нет, муж. (нем. Kraftmehl). Углевод особого состава, образующийся в виде мельчайших зернышек в зеленых частях растений из углекислоты воздуха под действием света (хим., бот.). || Продукт из таких зернышек различных растений … Толковый словарь Ушакова
КРАХМАЛ — муж. чисто мучнистая часть семян, особ. хлебных растение; добывается мочкою зерен, в виде белого порошка, более из пшеницы и картофеля; по клейкости своей, идет для придания жесткости и глади белыо, почему и называется также скорбилом (скорбнуть) … Толковый словарь Даля
Копрограмма
Копрограмма – это исследование кала (фекалий, экскрементов, стула), анализ его физических, химических свойств, а также разнообразных компонентов и включений различного происхождения. Она является частью диагностического исследования органов пищеварения и функции желудочно-кишечного тракта.
Синонимы русские
Общий анализ кала.
Синонимы английские
Koprogramma, Stool analysis.
Метод исследования
Микроскопия.
Какой биоматериал можно использовать для исследования?
Кал.
Как правильно подготовиться к исследованию?
Исключить прием слабительных препаратов, введение ректальных свечей, масел, ограничить прием медикаментов, влияющих на перистальтику кишечника (белладонны, пилокарпина и др.) и на окраску кала (железа, висмута, сернокислого бария), в течение 72 часов до сдачи кала.
Общая информация об исследовании
Копрограмма – это исследование кала (фекалий, экскрементов, стула), анализ его физических, химических свойств, а также разнообразных компонентов и включений различного происхождения. Она является частью диагностического исследования органов пищеварения и функции желудочно-кишечного тракта.
Кал – конечный продукт переваривания пищи в желудочно-кишечном тракте под воздействием пищеварительных ферментов, желчи, желудочного сока и жизнедеятельности кишечных бактерий.
По составу кал представляет собой воду, содержание которой в норме составляет 70-80 %, и сухой остаток. В свою очередь, сухой остаток на 50 % состоит из живых бактерий и на 50 % – из остатков переваренной пищи. Даже в пределах нормы состав кала в значительной степени непостоянен. Во многом он зависит от питания и употребления жидкости. Еще в большей степени состав кала варьируется при различных заболеваниях. Количество тех или иных компонентов в стуле изменяется при патологии или нарушении функции органов пищеварения, хотя отклонения в работе других систем организма также могут существенно влиять на деятельность желудочно-кишечного тракта, а значит, и на состав кала. Характер изменений при различных видах заболеваний чрезвычайно разнообразен. Можно выделить следующие группы нарушений состава фекалий:
- изменение количества компонентов, которые в норме содержатся в стуле,
- нерасщепленные и/или неусвоенные остатки пищи,
- биологические элементы и вещества, выделяемые из организма в просвет кишечника,
- различные вещества, которые образуются в просвете кишечника из продуктов обмена веществ, тканей и клеток тела,
- микроорганизмы,
- инородные включения биологического и другого происхождения.


Для чего используется исследование?
- Для диагностики различных заболеваний органов желудочно-кишечного тракта: патологии печени, желудка, поджелудочной железы, двенадцатиперстной, тонкой и толстой кишки, желчного пузыря и желчевыводящих путей.
- Для оценки результатов лечения заболеваний желудочно-кишечного тракта, требующих длительного врачебного наблюдения.
Когда назначается исследование?
- При симптомах какого-либо заболевания органов пищеварения: при болях в различных отделах живота, тошноте, рвоте, диарее или запоре, изменении окраски фекалий, крови в кале, ухудшении аппетита, потере массы тела, несмотря на удовлетворительное питание, ухудшении состояния кожи, волос и ногтей, желтушности кожи и/или белков глаз, повышенном газообразовании.
- Когда характер заболевания требует отслеживания результатов его лечения в процессе проводимой терапии.
Что означают результаты?
Референсные значения
|
Показатель |
Референсные значения |
|
Консистенция |
Плотная, оформленная, твердая, мягкая |
|
Форма |
Оформленный, цилиндрический |
|
Запах |
Каловый, кисловатый |
|
Цвет |
Светло-коричневый, коричневый, темно-коричневый, желтый, желто-зеленый, оливковый |
|
Реакция |
Нейтральная, слабокислотная |
|
Кровь |
Нет |
|
Слизь |
Отсутствует, небольшое количество |
|
Остатки непереваренной пищи |
Отсутствуют |
|
Мышечные волокна измененные |
Большое, умеренное, небольшое количество, отсутствуют |
|
Мышечные волокна неизмененные |
Отсутствуют |
|
Детрит |
Отсутствует, небольшое, умеренное, большое количество |
|
Растительная клетчатка переваримая |
Отсутствует, небольшое количество |
|
Жир нейтральный |
Отсутствует |
|
Жирные кислоты |
Отсутствуют, небольшое количество |
|
Мыла |
Отсутствуют, небольшое количество |
|
Крахмал внутриклеточный |
Отсутствует |
|
Крахмал внеклеточный |
Отсутствуют |
|
Лейкоциты |
Единичные в препарате |
|
Эритроциты |
0 — 1 |
|
Кристаллы |
Нет, холестерин, активуголь |
|
Йодофильная флора |
Отсутствует |
|
Клостридии |
Отсутствуют, небольшое количество |
|
Клетки кишечного эпителия |
Единичные в полях зрения или отсутствуют |
|
Дрожжеподобные грибы |
Отсутствуют |
Консистенция/форма
Консистенция стула определяется процентным содержанием в нем воды. Нормальным принято считать содержание воды в кале 75 %. В этом случае стул имеет умеренно плотную консистенцию и цилиндрическую форму, т. е. кал оформленный. Употребление повышенного объема растительной пищи, содержащей много клетчатки, приводит к усилению перистальтики кишечника, при этом кал становится кашицеобразным. Более жидкая консистенция, водянистая, связана с повышением содержания воды до 85 % и более.
Нормальным принято считать содержание воды в кале 75 %. В этом случае стул имеет умеренно плотную консистенцию и цилиндрическую форму, т. е. кал оформленный. Употребление повышенного объема растительной пищи, содержащей много клетчатки, приводит к усилению перистальтики кишечника, при этом кал становится кашицеобразным. Более жидкая консистенция, водянистая, связана с повышением содержания воды до 85 % и более.
Жидкий кашицеобразный кал называется диареей. Во многих случаях разжижение кала сопровождается увеличением его количества и частоты дефекаций в течение суток. По механизму развития диарею делят на вызываемую веществами, нарушающими всасывание воды из кишечника (осмотическую), возникающую в результате повышенной секреции жидкости из стенки кишечника (секреторную), являющуюся результатом усиления перистальтики кишечника (моторную) и смешанную.
Осмотическая диарея часто возникает в результате нарушения расщепления и усвоения элементов пищи (жиров, белков, углеводов). Изредка это может происходить при употреблении некоторых неусвояемых осмотически активных веществ (сульфата магния, соленой воды). Секреторная диарея является признаком воспаления кишечной стенки инфекционного и другого происхождения. Моторная диарея может вызываться некоторыми лекарственными веществами и нарушением функции нервной системы. Часто развитие того или иного заболевания связано с вовлечением как минимум двух механизмов возникновения диареи, такую диарею называют смешанной.
Изредка это может происходить при употреблении некоторых неусвояемых осмотически активных веществ (сульфата магния, соленой воды). Секреторная диарея является признаком воспаления кишечной стенки инфекционного и другого происхождения. Моторная диарея может вызываться некоторыми лекарственными веществами и нарушением функции нервной системы. Часто развитие того или иного заболевания связано с вовлечением как минимум двух механизмов возникновения диареи, такую диарею называют смешанной.
Твердый стул возникает при замедлении продвижения каловых масс по толстому кишечнику, что сопровождается их избыточным обезвоживанием (содержание воды в кале менее 50-60 %).
Запах
Обычный нерезкий запах кала связан с образованием летучих веществ, которые синтезируются в результате бактериальной ферментации белковых элементов пищи (индола, скатола, фенола, крезолов и др). Усиление этого запаха происходит при избыточном употреблении белковых продуктов или при недостаточном употреблении растительной пищи.
Резкий зловонный запах кала обусловлен усилением гнилостных процессов в кишечнике. Кислый запах возникает при усиленном брожении пищи, что может быть связано с ухудшением ферментативного расщепления углеводов или их усвоения, а также с инфекционными процессами.
Цвет
Нормальный цвет кала обусловлен присутствием в нем стеркобилина, конечного продукта обмена билирубина, который выделяется в кишечник с желчью. В свою очередь, билирубин является продуктом распада гемоглобина – основного функционального вещества красных клеток крови (гемоглобина). Таким образом, присутствие стеркобилина в кале – результат, с одной стороны, функционирования печени, а с другой – постоянного процесса обновления клеточного состава крови. Цвет кала в норме изменяется в зависимости от состава пищи. Более темный кал связан с употреблением мясной пищи, молочно-растительное питание приводит к осветлению стула.
Обесцвеченый кал (ахоличный) – признак отсутствия стеркобилина в стуле, к которому может приводить то, что желчь не попадает в кишечник из-за блокады желчевыводящих путей или резкого нарушения желчевыделительной функции печени.
Очень темный кал иногда является признаком повышения концентрации стеркобилина в стуле. В некоторых случаях это наблюдается при чрезмерном распаде эритроцитов, что вызывает усиленное выведение продуктов метаболизма гемоглобина.
Красный цвет кала может быть обусловлен кровотечением из нижних отделов кишечника.
Черный цвет – признак кровотечения из верхних отделов желудочно-кишечного тракта. В этом случае черная окраска стула – следствие окисления гемоглобина крови соляной кислотой желудочного сока.
Реакция
Реакция отражает кислотно-щелочные свойства стула. Кислая или щелочная реакция в кале обусловлена активизацией деятельности тех или других типов бактерий, что происходит при нарушении ферментации пищи. В норме реакция является нейтральной или слабощелочной. Щелочные свойства усиливаются при ухудшении ферментативного расщепления белков, что ускоряет их бактериальное разложение и приводит к образованию аммиака, имеющего щелочную реакцию.
Кислая реакция вызвана активизацией бактериального разложения углеводов в кишечнике (брожения).
Кровь
Кровь в кале появляется при кровотечении в желудочно-кишечном тракте.
Слизь
Слизь является продуктом выделения клеток, выстилающих внутреннюю поверхность кишечника (кишечного эпителия). Функция слизи заключается в защите клеток кишечника от повреждения. В норме в кале может присутствовать немного слизи. При воспалительных процессах в кишечнике усиливается продукция слизи и, соответственно, увеличивается ее количество в кале.
Детрит
Детрит – это мелкие частицы переваренной пищи и разрушенных бактериальных клеток. Бактериальные клетки могут быть разрушены в результате воспаления.
Остатки непереваренной пищи
Остатки пищи в стуле могут появляться при недостаточной продукции желудочного сока и/или пищеварительных ферментов, а также при ускорении перистальтики кишечника.
Мышечные волокна измененные
Измененные мышечные волокна – продукт переваривания мясной пищи.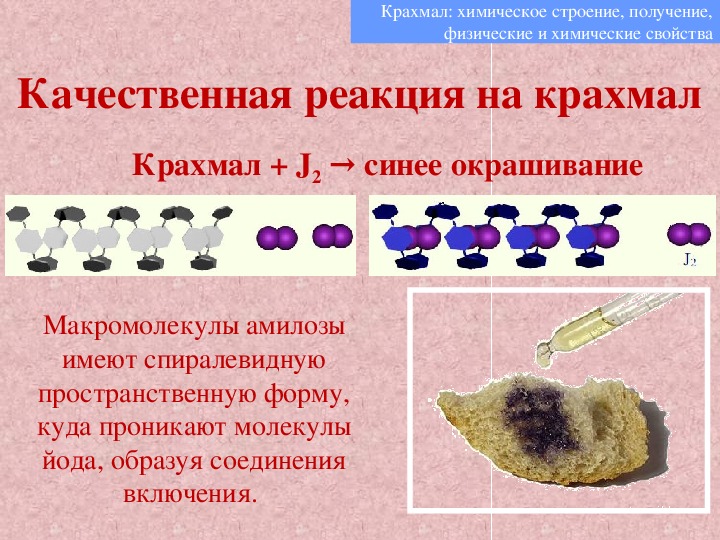 Увеличение содержания в кале слабоизмененных мышечных волокон происходит при ухудшении условий расщепления белка. Это может быть вызвано недостаточной продукцией желудочного сока, пищеварительных ферментов.
Увеличение содержания в кале слабоизмененных мышечных волокон происходит при ухудшении условий расщепления белка. Это может быть вызвано недостаточной продукцией желудочного сока, пищеварительных ферментов.
Мышечные волокна неизмененные
Неизмененные мышечные волокна – это элементы непереваренной мясной пищи. Их наличие в стуле является признаком нарушения расщепления белка (из-за нарушения секреторной функции желудка, поджелудочной железы или кишечника) либо ускоренного продвижения пищи по желудочно-кишечному тракту.
Растительная клетчатка переваримая
Переваримая растительная клетчатка – клетки мякоти плодов и другой растительной пищи. Она появляется в кале при нарушениях условий пищеварения: секреторной недостаточности желудка, усилении гнилостных процессов в кишечнике, недостаточном выделении желчи, нарушении пищеварения в тонком кишечнике.
Жир нейтральный
Нейтральный жир – это жировые компоненты пищи, которые не подверглись расщеплению и усвоению и поэтому выводятся из кишечника в неизменном виде. Для нормального расщепления жира необходимы ферменты поджелудочной железы и достаточное количество желчи, функция которой заключается в разделении жировой массы на мелкокапельный раствор (эмульсию) и многократном увеличении площади соприкосновения жировых частиц с молекулами специфических ферментов – липаз. Таким образом, появление нейтрального жира в кале является признаком недостаточности функции поджелудочной железы, печени или нарушения выделения желчи в просвет кишечника.
Для нормального расщепления жира необходимы ферменты поджелудочной железы и достаточное количество желчи, функция которой заключается в разделении жировой массы на мелкокапельный раствор (эмульсию) и многократном увеличении площади соприкосновения жировых частиц с молекулами специфических ферментов – липаз. Таким образом, появление нейтрального жира в кале является признаком недостаточности функции поджелудочной железы, печени или нарушения выделения желчи в просвет кишечника.
У детей небольшое количество жира в кале может являться нормой. Это связано с тем, что органы пищеварения у них еще недостаточно развиты и поэтому не всегда справляются с нагрузкой по усвоению взрослой пищи.
Жирные кислоты
Жирные кислоты – продукты расщепления жиров пищеварительными ферментами – липазами. Появление жирных кислот в стуле является признаком нарушения их усвоения в кишечнике. Это может быть вызвано нарушением всасывательной функции кишечной стенки (в результате воспалительного процесса) и/или усилением перистальтики.
Мыла
Мыла – это видоизмененные остатки неусвоенных жиров. В норме в процессе пищеварения усваивается 90-98 % жиров, оставшаяся часть может связываться с солями кальция и магния, которые содержатся в питьевой воде, и образовывать нерастворимые частицы. Повышение количества мыл в стуле является признаком нарушения расщепления жиров в результате недостатка пищеварительных ферментов и желчи.
Крахмал внутриклеточный
Внутриклеточный крахмал – это крахмал, заключенный внутри оболочек растительных клеток. Он не должен определяться в кале, так как при нормальном пищеварении тонкие клеточные оболочки разрушаются пищеварительными ферментами, после чего их содержимое расщепляется и усваивается. Появление внутриклеточного крахмала в кале – признак нарушения пищеварения в желудке в результате уменьшения секреции желудочного сока, нарушения пищеварения в кишечнике в случае усиления гнилостных или бродильных процессов.
Крахмал внеклеточный
Внеклеточный крахмал – непереваренные зерна крахмала из разрушенных растительных клеток. В норме крахмал полностью расщепляется пищеварительными ферментами и усваивается за время прохождения пищи по желудочно-кишечному тракту, так что в кале не присутствует. Появление его в стуле указывает на недостаточную активность специфических ферментов, которые ответственны за его расщепление (амилаза) или слишком быстрое продвижение пищи по кишечнику.
В норме крахмал полностью расщепляется пищеварительными ферментами и усваивается за время прохождения пищи по желудочно-кишечному тракту, так что в кале не присутствует. Появление его в стуле указывает на недостаточную активность специфических ферментов, которые ответственны за его расщепление (амилаза) или слишком быстрое продвижение пищи по кишечнику.
Лейкоциты
Лейкоциты – это клетки крови, которые защищают организм от инфекций. Они накапливаются в тканях тела и его полостях, там, где возникает воспалительный процесс. Большое количество лейкоцитов в кале свидетельствует о воспалении в различных отделах кишечника, вызванном развитием инфекции или другими причинами.
Эритроциты
Эритроциты – красные клетки крови. Число эритроцитов в кале может повышаться в результате кровотечения из стенки толстого кишечника или прямой кишки.
Кристаллы
Кристаллы образуются из различных химических веществ, которые появляются в кале в результате нарушения пищеварения или различных заболеваний. К ним относятся:
- трипельфосфаты – образуются в кишечнике в резкощелочной среде, которая может являться результатом активности гнилостных бактерий,
- гематоидин – продукт превращения гемоглобина, признак выделения крови из стенки тонкого кишечника,
- кристаллы Шарко – Лейдена – продукт кристаллизации белка эозинофилов – клеток крови, которые принимают активное участие в различных аллергических процессах, являются признаком аллергического процесса в кишечнике, который могут вызывать кишечные гельминты.
Йодофильная флора
Йодофильной флорой называется совокупность различных видов бактерий, которые вызывают бродильные процессы в кишечнике. При лабораторном исследовании они могут окрашиваться раствором йода. Появление йодофильной флоры в стуле является признаком бродильной диспепсии.
Клостридии
Клостридии – разновидность бактерий, которые могут вызывать в кишечнике гниение. Увеличение числа клостридий в стуле указывает на усиление гниения в кишечнике белковых веществ вследствие недостаточной ферментации пищи в желудке или кишечнике.
Эпителий
Эпителий – это клетки внутренней оболочки кишечной стенки. Появление большого числа эпителиальных клеток в стуле является признаком воспалительного процесса кишечной стенки.
Дрожжеподобные грибы
Дрожжеподобные грибы – разновидность инфекции, которая развивается в кишечнике при недостаточной активности нормальных кишечных бактерий, препятствующих ее возникновению. Их активное размножение в кишечнике может быть результатом гибели нормальных кишечных бактерий из-за лечения антибиотиками или некоторыми другими лекарственными средствами. Кроме того, появление грибковой инфекции в кишечнике иногда является признаком резкого снижения иммунитета.
С картошкой все не так просто – Наука – Коммерсантъ
Систематика
Отдел Цветковые Angiospermae
Класс Двудольные Magnoliopsida
Порядок Пасленоцветные Solanales
Семейство Пасленовые Solanaceae
Род Паслен Solanum
Подсекция potatoe
Вид Solanum tuberosum
Подвид tuberosum
Картофель — многолетнее травянистое растение из семейства пасленовых. Его плоды ядовиты, в пищу употребляют только клубни — видоизмененные подземные побеги. То, что мы называем картофелем, содержит лишь малую толику его настоящего генетического разнообразия. В Южной Америке растет около 200 его диких видов и подвидов, но все известные сейчас культурные сорта (более 5000) относятся к одному роду Solanum. Этот род подразделяется на несколько подсекций, и все клубнеобразующие виды картофеля относятся к подсекции potato; сейчас считается, что весь культивируемый картофель относится к 4 видам (раньше его разделяли на 7-10): это Solanum tuberosum и три гибридных вида горького картофеля. S.tuberosum подразделяется на два подвида: tuberosum и andigena. Первый представляет собой тот самый картофель, который едят сейчас во всем мире, второй — сельскохозяйственные культуры, которые ограниченно выращивают только в Центральной и Южной Америке.
История вопроса
Родина картофеля — Южная Америка, а точнее, перуанские Анды. Долгое время считалось, что картофель был введен в культуру одновременно в нескольких местах, что ранние культивируемые формы имеют независимое происхождение от нескольких разных видов, но последние генетические исследования показали, что это не так. Одомашнивание картофеля произошло в высокогорьях южного Перу около 7-10 тыс. лет назад, после чего картофель стал основным продуктом питания инков. В Перу существуют сотни его местных разновидностей, когда-то практически каждая семья выращивала свой собственный сорт картофеля, и знания по выращиванию этих уникальных клубней передавались из поколения в поколение. Крестьяне постоянно проводили селекцию новых и новых сортов, добиваясь устойчивости к разнообразным вредителям, болезням и климатическими изменениями. Благодаря этому количество сортов картофеля быстро увеличивалось, обеспечивая очень высокое биоразнообразие. Будучи основной пищей андских крестьян, картофель также играл и важнейшую роль в их культуре: единицей измерения времени у них был период, необходимый для приготовления горшка картофеля, а единицей измерения площади служил участок (топо), с которого можно собрать достаточный для прокорма одной семьи в течение сезона урожай картофеля.
В Европе картофель появился во второй половине XVI века после испанского завоевания инков. Из Нового Света в Европу прибыла лишь небольшая часть андского многообразия, причем далеко не самая удачная — неустойчивая к фитофторозу, колорадскому жуку и нематоде. Ограниченное генетическое разнообразие у завезенного в Европу картофеля привело к постепенному вырождению этой культуры и сделало ее очень уязвимой: паразиты и болезни быстро распространялись с одного растения на другие. В 1840-х годах по Европе прошла эпидемия картофельного фитофтороза, от которой больше всех пострадала Ирландия, где картофель к тому времени успел стать основным продуктом питания (на него приходилось 80% потребляемых калорий).
Сорт картофеля «Сирень»
Фото: ГНУ ВНИИКХ им. А.Г. Лорха
В России картофель стали выращивать при Петре I, но лишь с целью использовать его как лекарственное растение. Всерьез заниматься картофелем начала лишь Екатерина II. Она поручила начать его разведение Абраму Ганнибалу, уже имевшему с картофелем дело. Вскоре Екатерина приказала разослать клубни картофеля вместе с инструкциями по его разведению по губерниям. Но крестьяне не желали принимать новую культуру (тем более что ей приписывали дьявольские свойства) и встретили ее картофельными бунтами. Тем не менее, с 1840 года площади картофельных полей в России начали интенсивно увеличиваться, и уже через несколько десятилетий картофель не только признали в народе, но и стали называть «вторым хлебом».
Теперь картофель выращивают почти в 100 странах, это четвертая продовольственная культура в мире — после риса, пшеницы и кукурузы. Он успешно растет в умеренных, субтропических и тропических широтах, предпочитая при этом прохладную погоду: при температуре ниже 10°C и выше 30°C рост клубней резко замедляется. В тропическом климате картофель растет в холодные месяцы года.
Сейчас больше всего картофеля на душу населения производят в Европе (особенно в Восточной и Центральной Европе), но ей в затылок уже дышит южная и восточная Азия. Китай уже сейчас выращивает самые большие урожаи картофеля в мире, на 2 месте — Россия, которая собирает вдвое меньше, 3 место у Индии. В России средняя урожайность картофеля 13 т/га, в Китае — 14,5 т/га, а, например, в Голландии — 45 т/га. Россия значительно отстает даже от среднего мирового уровня (17 т/га). В год у нас в стране собирают около 30 млн тонн, из них около 100 тыс. тонн экспортируется за рубеж, в то время как импортируется около 500 тысяч тонн.
Зачем нужна картошка
Картофель выращивается не только в строго пищевых целях. Его используют в качестве корма для домашних животных, для производства алкогольных напитков. Картофельный крахмал может применяться в пищевой промышленности как загуститель для супов и соусов, в текстильной промышленности, а также для изготовления клея, бумаги и картона. Сейчас изучается возможность использования отходов картофеля для получения полимолочной кислоты, применяемой в производстве пластмассовых изделий; ведутся исследовательские работы по поиску способов использования крахмала в качестве основы для экологически чистой упаковки.
Холодная картошка полезнее
Годовой рацион современного человека составляет около 33 кг картофеля. Средних размеров картофелина весит 150 г и содержит примерно 27 мг витамина С (45% от дневной нормы), 620 мг калия (18% от дневной нормы), 0,2 мг витамина В6 (10% от дневной нормы), а также тиамин, рибофлавин, фолиевую кислоту, ниацин, магний, фосфор, железо и цинк. Картофель известен высоким содержанием углеводов (примерно 26 г в картофелине среднего размера). Преобладающей формой углеводов в картофеле является крахмал (в среднем 17,5% в свежем картофеле или 75-80% в пересчете на сухое вещество).
Картофельный крахмал состоит из разветвленного амилопектина и линейной амилозы, их соотношение зависит от сорта картофеля. Амилоза, с длинными цепями молекул, водорастворима, она диффундирует из гранул крахмала при варке в воде. Амилопектин с сильно разветвленными молекулами состоит из той же амилозы и более сложного углевода — пектина. Сорта с более высоким содержанием амилопектина меньше развариваются и сохраняют свою форму при варке. Небольшая часть этого крахмала устойчива к перевариванию и не всасывается в тонком кишечнике — это резистентный крахмал. Причем чем больше содержание амилозы, тем выше будет доля резистентных крахмалов в картофеле. Считается, что этот крахмал — так же, как волокна целлюлозы — обеспечивает защиту толстой кишки от рака, снижает уровень холестерина в крови, повышает ощущение сытости и даже снижает накопление жира. Количество резистентного крахмала в картофеле во многом зависит от способа его приготовления. Например, если в готовом картофеле содержится около 7% резистентного крахмала, то при охлаждении его становится больше примерно на 13%.
Не отравишься, так заболеешь
Как и другие представители семейства пасленовых (белена, дурман, табак, паслен), картофель содержит токсичные соединения — гликоалкалоиды, из которых наиболее распространенны соланин и чаконин. Соланин обладает фунгицидными и инсектицидными свойствами и защищает растение от хищников. Для человека и животных он токсичен даже в небольших дозах. Как правило, ядовитые соединения накапливаются в листьях, стеблях, побегах и плодах, но воздействие света и физические повреждения приводят к повышению содержания гликоалкалоидов в клубнях. Особенно много гликоалкалоидов сосредоточено непосредственно под кожей, в позеленевших и проросших клубнях. В диком картофеле концентрация токсинов достаточно высока для отравления человека. Они угнетающе действуют на центральную нервную систему, могут вызывать головную боль, диарею, обезвоживание, лихорадку, судороги, а в тяжелых случаях кому и смерть, однако, в реальности отравления картофелем происходят очень редко. Некоторые сорта картофеля отличаются особенно высоким содержанием гликоалкалоидов; от них селекционерам приходится отказываться, даже если они перспективны в других отношениях.
Профессор Виктор Старовойтов, заместитель директора по науке ВНИИ картофельного хозяйства имени А.Г. Лорха, доктор технических наук
Фото: Евгений Дудин, Коммерсантъ
«Содержание алкалоидов в мякоти клубней составляет 10-50 мг/кг, — рассказывает замдиректора по науке ВНИИ картофельного хозяйства имени Лорха профессор Виктор Старовойтов, — в кожице клубня их концентрация выше; в целом в клубне их содержится от 20 до 100 мг/кг. При высоком содержании алкалоидов (150-200 мг/кг сырого веса) картофель становится горьким, при концентрации 230-270 мг алкалоидов на килограмм сырого веса может наступить отравление организма. Однако при кулинарной обработке количество гликоалкалоидов в клубнях снижается за счет термического разрушения до более простых соединений, которые, к тому же, частично вымываются и остаются в растворе».
В 2002 году в картофеле было обнаружено еще одно вредное вещество — акриламид. Акриламид появляется при высокотемпературной обработке многих крахмалистых продуктов (в первую очередь, картофеля и злаков), если их жарить, готовить во фритюре, запекать в духовке или на гриле. В тех же самых сырых или вареных продуктах его нет. Дальнейшие исследования показали, что причина не в крахмале: картофель и злаковые наряду с крахмалом содержат аминокислоту аспарагин, а при нагревании от 120 градусов и выше при взаимодействии с сахарами аспарагин превращается в акриламид. Если же продукты не жарить, а варить, акриламид в них не образуется совсем или его содержание незначительно.
«Еще 10 лет назад никто не знал о том, что акриламид может содержаться в пищевых продуктах, — говорит Софья Лущеницкая, научный сотрудник химфака МГУ, — было известно, что это вещество может содержаться в пластиковых упаковках, в табачном дыме, иногда в небольшом количестве попадает в воду, и что оно обладает мутагенными свойствами. И вдруг в 2002 году ученые из Стокгольмского университета обнаружили, что во многих продуктах содержание акриламида в сотни и тысячи раз превышает любые возможные ПДК. Он в разных концентрациях был обнаружен в чипсах, жареном картофеле, в хрустящих хлебцах, выпечке, мюслях и кукурузных хлопьях». Авторы работы, опубликованной в 2009 году в American Journal of Clinical Nutrition, обнаружили, что если ежедневно в течение двух недель потреблять 157 мг акриламида из картофельных чипсов, это может запустить процессы, которые в итоге становятся причиной заболеваний сердца. У женщин, часто употребляющих акриламидные продукты, рак груди встречается в 2 раза чаще, чем у тех, кто их избегает. Есть данные, что это вещество увеличивает риск рака яичников на 79%, матки — на 28%, а почек — на 59%.
Генетика и селекция
Во Всероссийском научно-исследовательском институте картофельного хозяйства имени А.Г. Лорха безвирусный семенной картофель выращивают методом микроклонального размножения in vitro
Фото: Евгений Дудин, Коммерсантъ
В 2009 году была закончена расшифровка генома картофеля. Картофельный геном имеет средний для растений размер, он содержит 12 хромосом и 860 млн пар оснований. Расшифровкой каждой хромосомы картофеля занималось одно или несколько государств; российские ученые из Центра биоинженерии РАН приняли участие в расшифровке 12-й хромосомы. Картофель S. tuberosum tuberosum представлен диплоидомыми (2n=24) или тетраплоидомыми (4n=48) формами. Диплоиды встречаются только в Чили, а культивируемый во всем мире картофель является тетраплоидом. Происхождение тетраплоидов обеспечивается благодаря характерному для видов рода Solanum феномену нередуцированных гамет: у большинства из них, кроме нормальных гаплоидных гамет, с частотой 2-10% могут встречаться гаметы с нередуцированным (двойным) числом хромосом.
Для того чтобы повысить резистентность картофеля к вредителям и болезням, повысить урожайность и усилить его ценные качества, люди уже много веков подряд занимаются селекцией картофеля. А так как картофель размножается в основном вегетативным путем, селекционеры не отбирают его по способности цветков привлекать опылителей. В результате большинство сортов картофеля имеют пониженную способность к цветению, а естественное — перекрестное — опыление необходимо для поддержания разнообразия. Цветущие сорта картофеля, способные привлекать опылителей, еще остались в мелких фермерских хозяйствах в Андах, но и там многие древние сорта, традиционно выращиваемые перуанскими фермерами, уже утрачены. Для сохранения и защиты того, что осталось, в Перу создан крупнейший в мире банк, где в стерильных условиях хранятся образцы генетического материала — около 100 диких видов картофеля из восьми латиноамериканских стран, а также 3800 традиционных андских сортов.
Генная модификация
В начале 1990-х годов транснациональная корпорация Monsanto, лидер в сфере высоких технологий в сельском хозяйстве, выпустила для коммерческого использования трансгенные сорта картофеля, резистентные к колорадскому жуку и вирусным заболеваниям. Устойчивость к вредителям достигалась благодаря включению генов бактерии Bacillus thuringiensis, которая вырабатывает специфический белковый эндоксин, обладающий инсектицидным действием. Немецкая химическая компания BASF создала модифицированный картофель (сорт Amflora), состав которого был изменен (он содержит только амилопектин) так, что он стал несъедобным, но более подходящим для производства крахмала. После долгих споров Еврокомиссия в прошлом году разрешила выращивать этот сорт в ЕС в промышленных целях. В прошлом же году группа индийских ученых объявила, что они разработали генетически модифицированный картофель, который содержит вдвое больше белка, чем обычный (благодаря добавлению гена AmA1 амаранта).
Трансгенные сорта картофеля позволяют значительно увеличивать его урожайность, добиться появления принципиально новых свойств и открывают большие возможности для его использования в промышленности. Но общество с недоверием относится к генномодифицированным продуктам, и многие крупные компании (McDonald’s, Burger King, Frito-Lay, и Procter&Gamble) отказываются от использования ГМ-картофеля. Впрочем, ученые надеются, что развивающиеся страны, где более миллиарда человек страдает от хронического недоедания, отнесутся к нему с большим интересом.
ГМ-картофель в России
В России с генномодифицированным картофелем ситуация странная. У нас сертифицировано 4 сорта ГМ-картофеля, два из которых разработала Monsanto, а два («Елизавета плюс» и «Луговской плюс») — российский Центр биоинженерии РАН, но сертифицированы они только для употребления в пищу: есть их в России можно, а выращивать — нельзя.
Россия является одной из главных картофелеводческих стран мира, но из-за колорадского жука происходят огромные потери урожая, особенно в южных регионах страны. Ежегодные потери от колорадского жука оцениваются в 2-2,5 млрд долларов. Генномодифицированный картофель, устойчивый к нему, мог бы решить эту проблему. Он прост в выращивании и не требует применения инсектицидов. «Мы разработали 2 сорта генномодифицированного картофеля, устойчивого к колорадскому жуку, — рассказывает доктор биологических наук, замдиректора по научной работе Центра биоинженерии РАН Николай Равин. — Эти сорта жук не ест. Мы проводили полевые испытания: высаживали две грядки картофеля, на одной наш сорт (устойчивый), на другой — исходный, от которого он произошел. В результате одна грядка оказывалась полностью съедена жуком, другая же осталась неповрежденной. Существуют еще американские сорта (созданные Monsanto), но они не приспособлены к нашим климатическим условиям. Мы же брали сорта, которые давно и успешно выращиваются в России и привносили в них дополнительный признак — устойчивость к колорадскому жуку».
«На данный момент у нас нет разрешения продавать семенной материал для выращивания на полях: есть можно, импортировать можно, а выращивать нельзя, — продолжает Равин. — Люди боятся генномодифицированных продуктов, на мой взгляд, совершенно безосновательно. Им по телевизору рассказывают страшилки, что взяли ген какого-нибудь страшного организма (например, скорпиона), вставили в картошку, и если вы съедите эту картошку, то сами превратитесь в скорпиона. Научной основы под этими страшилками нет никакой. Другой вопрос, что существует опасность утечки модифицированного генного материала в окружающую среду — опасность передачи новых генов диким родственникам. Поэтому генномодифицированные сорта должны сажаться отдельно, нужно избегать их контакта с сородичами; но это уже вопрос техники ведения сельского хозяйства. В случае картофеля это проблема вообще не очень актуальна: размножается он, в основном, вегетативным путем».
Татьяна Вайнтроб
Крахмал кукурузный — описание, состав, калорийность и пищевая ценность
Мелкий сыпучий порошок белого или светло-желтого цвета, который может набухать в горячей или холодной воде, не меняя химических свойств.
Изготовление
Чтобы получить крахмал, зерна кукурузы замачиваются в растворе сернистой кислоты. Это приводит к тому, что белок, который связывает крахмал, растворяется. Затем зерно дробят, что приводит к выделению крахмального молока. Процесс отделения крахмала от белка, который не растворился, происходит в центрифугах. Готовый продукт промывают, высушивают и упаковывают.
Калорийность
В 100 граммах продукта содержится 340 кКал.
Состав
Кукурузный крахмал с основном состоит из сложных углеводов. Также в его состав входят зола, незначительное количество клетчатки, витамина PP и таких минеральных элементов, как натрий, кальций, калий, фосфор, магний.
Использование
Кукурузный крахмал широко применяется в кулинарии для изготовления разнообразных блюд, в том числе супов, соусов, лепешек, хлеба, блинов, маффинов, кексов, кляра.
При выпечке его можно использовать в качестве заменителя пшеничной муки, однако общий объем крахмала не должен превышать 50%.
В качестве загустителя кукурузный крахмал выгодно отличается от муки тем, что практически не влияет на вкус конечного блюда, не содержит глютен, действует более эффективно, делает блюдо прозрачным, блестящим. Для сгущения 375-500 мл жидкости требуется одна столовая ложка крахмала.
Его используют не только в кулинарии, но и в фармакологической промышленности для изготовления присыпок, мазей, микстур, а также в косметологии при производстве паст, пудр.
Полезные свойства
О полезности кукурузного крахмала идут многочисленные споры. Он представляет собой сложный углевод, поэтому медленно растворяется и не приводит к скачкам сахара в крови. Кроме того, крахмал помогает поддерживать работу иммунной системы, участвует в образовании органических кислот и мышечной массы, препятствует появлению воспалительных процессов, питает нервные клетки.
Ограничения по употреблению
Кукурузный крахмал содержит глюкозу и сахара, что может привести к увеличению веса. Однако для этого его нужно употреблять регулярно и в большом количестве.
Редко может проявляться индивидуальная непереносимость крахмала.
Его не рекомендуют употреблять при изжоге, гастрите, ожирении, болезнях желудочно-кишечного тракта.
Волшебник «синий йод»
Это йодированный крахмал. Его легко приготовить в домашних условиях. Для этого надо в 50 мл теплой воды развести 10 г картофельного крахмала (1 ч. ложка с верхом), размешать, добавить 10 г сахарного песка и 0,4 г лимонной кислоты (несколько кристалликов). Одновременно с этим вскипятить 150 мл воды и в кипяток влить полученный раствор крахмала. Смесь размешать, охладить и добавить 1 ч. ложку 5-процентного спиртового раствора йода, после чего она окрасится в синий цвет. Это и есть тот самый «синий йод».
Присутствующие в препарате сахар и лимонная кислота необходимы для улучшения его вкуса. Кроме того, они препятствуют разложению «синего йода», поэтому он может храниться в закрытом сосуде при комнатной температуре, не теряя своих свойств, в течение многих месяцев.
«Синий йод» открывали несколько раз. Но наиболее полное описание его лечебных свойств дал ученый из Санкт-Петербурга В. Мохнач. Он впервые опробовал этот препарат на себе в годы Великой Отечественной войны, когда заболел тяжелой формой бактериальной дизентерии. А в 50-х годах с его участием в медицинских учреждениях Ленинграда было проверено действие препарата на большой группе больных дизентерией, колитами и энтероколитами, состояние которых оценивалось как тяжелое и средней тяжести. Суточная доза приема препарата для этих больных составляла 500 г. Нередко бывает, что при лечении острых и хронических кишечных инфекций применяются и значительно большие дозы. Достаточно сказать, что сам Мохнач увеличивал дозу приема «синего йода» до 1500–2000 г в сутки.
При язвенных поражениях толстого кишечника «синий йод» вводится в виде клизм по 50 г ежедневно в течение недели.
При конъюнктивите в течение недели в глаза закапывается по 2–3 капли специально приготовленного раствора (1 ч. ложка «синего йода» разводится в 10 ч. ложках теплой дистиллированной воды) утром и вечером.
При стоматите препарат разводят теплой водой и используют этот раствор для полоскания полости рта 2–3 раза в день.
При приступах колита чайными ложками пейте «синий йод». В начале лечения может наблюдаться рвота, не бойтесь этого. Если приступ колита начали лечить вечером, то до утра следует выпить не менее стакана (суточная норма). В дальнейшем лечение можно продолжать, пока болезнь не утихнет совсем.
Расстройства желудка. Известная российская целительница В. Травинка лечит этот недуг комбинацией «синего йода» и голубой целительной глины. Предварительно готовит глиняную воду (1-1,5 столовой ложки глины на стакан воды) и дает выпить этот раствор за час до приема «синего йода». Глина обладает прекрасным свойством вытягивать шлаки и токсины из организма, а йод обеззараживает и нейтрализует организм от вредных микробов.
Ожоги. Налейте «синий йод» в пластиковую бутылку любой емкости с резьбой и через пульверизатор — чтобы щадить рану- распыляйте лекарство по обожженному месту. Можно наложить на обожженное место смоченный в «синем йоде» тройной слой марли. По мере высыхания компресса снова смочите марлю.
С помощью «синего йода» улучшается эластичность кровеносных сосудов, что позволяет рекомендовать его для лечения последствий инсульта. «Синий йод» помогает нормализовать и высокое, и низкое кровяное давление. Кроме того, благодаря седативным свойствам его можно принимать в качестве успокоительного средства.
Соединение йода с крахмалом нейтрализует его токсичные свойства, поэтому «синий йод» не вызывает отравления и может употребляться в значительных дозах. Кроме того, входящий в состав препарата крахмал при приеме внутрь обволакивает язвы и другие пораженные места слизистой желудочно-кишечного тракта, создавая некий защитный слой, способствующий быстрому излечению. Сам же йод стимулирует деятельность поджелудочной железы, являющейся защитницей всего нашего организма.
Щитовидная железа не только защищает наш организм от вирусов и микробов, но и активно участвует в обмене веществ, снимает нервное напряжение, пополняет энергетические ресурсы организма, потраченные нами в течение трудового дня. Нарушение же функций щитовидной железы приводит организм к расстройству, так как щитовидной железе подвластны и центральная нервная система, и процессы кроветворения, и сопротивляемость организма инфекциям и даже раковым клеткам.
А нормально щитовидная железа может функционировать только при достаточном количестве йода в организме. Это примерно 300 мг в сутки. Поэтому наряду с лечением тех или иных болезней «синий йод» рекомендуется принимать и для профилактики йодной недостаточности. Продолжительность профилактического курса составляет не более месяца. В этот период «синий йод» следует принимать 2 раза в неделю по 1 ч. ложке. Для тех, у кого вес превышает 65 кг, дозу приема можно увеличить до 2 ч. ложек.
Есть еще аптечный препарат «йодинол», приготовленный на основе поливинилового спирта. «Йодинол» — это тоже «синий йод». Он является прекрасным антисептиком, нетоксичный, недорогой и очень устойчивый препарат, который может долго храниться.
Необходимую организму дозу йода можно ввести также с помощью раствора Люголя. Доза приема — всего одна капля. Ее рекомендуется принимать каждую неделю по вторникам и пятницам до еды. Для этого в стакан с подкисленной яблочным уксусом теплой кипяченой водой надо добавить одну каплю раствора Люголя, тщательно перемешать и выпить весь стакан этой смеси небольшими глотками, запивая чаем или кофе. В условиях повышенной нагрузки на организм эту процедуру надо выполнять ежедневно до тех пор, пока нагрузки на организм не уменьшатся.
Первым признаком йодной недостаточности в организме является сильная усталость, быстрая утомляемость, повышенная раздражительность, чувство разбитости по утрам. При появлении этих тревожных сигналов вспомните о «синем йоде». С его помощью вам удастся укрепить здоровье и сохранить хорошее настроение. Он даст вашему организму такой заряд бодрости, что вы без труда сможете отказаться от кофе и крепкого чая.
«Синий йод» сохраняет свои лечебные свойства до тех пор, пока сохраняется его темный цвет. Единственное ограничение – его не следует принимать людям, страдающим тромбофлебитом.
Татьяна ПЕТРОВА.
Диетические полисахариды
Диетические полисахариды
Полисахариды, особенно растительного происхождения, являются важными компонентами в рационах травоядных и всеядных животных. Этот сложный набор молекул можно разделить на несколько категорий, в зависимости от того, идет ли речь о химии или питании. С точки зрения физиологии пищеварения и питания, возможно, наиболее подходящая классификация основана на том, синтезируют ли животные ферменты, которые позволяют рассматриваемому полисахариду расщепляться до абсорбируемых моносахаридов.С этой точки зрения, у нас есть крахмал, который может перевариваться ферментами позвоночных, а не клетчатка, которая не может.
Крахмал: амилоза и амилопектин
Крахмал — это основной углевод, содержащийся в семенах и клубнях растений; важные источники крахмала включают кукурузу (кукурузу), картофель и рис. Крахмал существует в виде гранул, каждая из которых состоит из нескольких миллионов молекул амилопектина вместе с еще большим количеством молекул амилозы. Поскольку амилопектин представляет собой гораздо более крупную молекулу, чем амилоза, масса амилопектина обычно в 4-5 раз превышает массу амилозы в крахмале.
| Амилоза состоит из линейных спиральных цепей примерно от 500 до 20 000 мономеров альфа-D-глюкозы, связанных вместе альфа (1-4) гликозидными связями. | Молекулы амилопектина — это огромные разветвленные полимеры глюкозы, каждый из которых содержит от одного до двух миллионов остатков. В отличие от амилозы амилопектин является разветвленным. Он содержит многочисленные амилозоподобные цепи до 30 остатков глюкозы, связанных альфа (1-4) связями, соединенных друг с другом через точки разветвления альфа (1-6). |
Крахмал расщепляется до глюкозы в два основных этапа:
Сначала амилоза и амилопектин гидролизуются на мелкие фрагменты под действием альфа-амилазы, секретируемой слюнными железами у некоторых видов и поджелудочной железой в целом. Амилаза расщепляет только внутренние альфа (1-4) гликозидные связи, тем самым восстанавливая крахмал до трех различных олигосахаридов: мальтозы (дисахарид), мальтотриозы (трисахарида) и группы декстринов с ограничением альфа, которые содержат точки ветвления от амилопектина.
Во-вторых, мальтоза, мальтотриоза и предельные декстрины гидролизуются на просветной поверхности тонкой кишки ферментным комплексом щеточной каймы, называемым сукраза-изомальтаза (также часто называемым мальтазой). На этом этапе в конечном итоге получают мономеры глюкозы, которые затем транспортируются в энтероциты тонкого кишечника за счет совместного транспорта с ионами натрия.
Пищевые волокна: целлюлоза и гемицеллюлоза
Было предложено несколько определений для «волокна». Раннее определение, все еще вполне подходящее, в основном гласит, что клетчатка — это часть пищи, полученная из стенок растительных клеток, которая плохо переваривается млекопитающими.«Другими словами, млекопитающие часто потребляют клетчатку, но сами не выделяют ферменты, необходимые для ее переваривания в форму, которая может усваиваться. Другое общее определение клетчатки — это некрахмальный полисахаридный компонент пищевых продуктов.
Основными компонентами пищевых волокон являются целлюлоза и гемицеллюлоза растительного происхождения. Пектин и пектиновая кислота — это другие полисахариды растений, часто присутствующие в диетах.
Целлюлоза представляет собой линейный полимер, содержащий от 1000 до 10000 молекул бета-D-глюкозы, в котором соседние молекулы глюкозы ковалентно связаны через бета (1-4) гликозидные связи.Бета (1-4) связи заставляют полимер принимать неспиральную, прямую структуру, которая отличается от спиральной структуры, наложенной на молекулы крахмала альфа (1-4) связью. Не спиральная структура целлюлозы также способствует образованию водородных связей между молекулами целлюлозы.
Полимеры целлюлозы связываются друг с другом посредством огромного количества водородных связей, образуя микроволокна. Микроволокна взаимодействуют с образованием целлюлозных волокон. Типичное волокно содержит примерно 500 000 молекул целлюлозы.Высокая прочность на разрыв целлюлозных волокон отражает огромное количество водородных связей, включенных в их структуру.
Гемицеллюлоза представляет собой гетерополимер, состоящий из различных сахаров, включая ксилозу, арабинозу и маннозу, с разветвленной структурой. В отличие от высокоупорядоченной структуры целлюлозы, гемицеллюлоза принимает аморфозную структуру и становится сильно гидратированной, образуя гель.
Клетка позвоночных, вырабатывающая фермент, гидролизующий целлюлозу или гемицеллюлозу, не идентифицирована.Конечно, амилаза не расщепляет эти два полисахарида. Таким образом, пищевые волокна не перевариваются и проходят через тонкий кишечник практически без изменений.
В толстой кишке (или в предсердиях жвачных животных) клетчатка переваривается ферментами — целлюлазами и гемецеллюлазами — микробного происхождения; этот процесс называется ферментацией. Ферментация не дает моносахаридов, которые можно абсорбировать. Скорее, его основными продуктами являются летучие жирные кислоты с короткой цепью, которые легко абсорбируются и используются для получения энергии и синтеза липидов.Таким образом, если чан для брожения имеет достаточный размер (например, для травоядных животных), пищевые волокна могут быть основным источником энергии. 11
Отправляйте комментарии [email protected]
Образование крахмала в клетках растений
Реферат
Богатые крахмалом культуры составляют основу нашего питания, но растениям еще предстоит раскрыть все свои секреты того, как они производят это жизненно важное вещество. Большой прогресс был достигнут в изучении как сельскохозяйственных культур, так и модельных систем, и мы приближаемся к пониманию ферментативного механизма, ответственного за создание массивных нерастворимых гранул крахмала, обнаруженных в тканях растений.Здесь мы суммируем наше текущее понимание этих биосинтетических ферментов, подчеркивая недавний прогресс в выяснении их конкретных функций. Тем не менее, во многих отношениях мы только прикоснулись к поверхности: остается много неуверенности в том, как эти компоненты работают вместе и контролируются. Мы отмечаем недавние наблюдения, предполагающие значительную степень гибкости во время синтеза крахмала и то, что ранее не подозреваемые неферментативные белки могут иметь значение. Мы пришли к выводу, что исследование крахмала еще не является зрелой темой, и что новые экспериментальные и теоретические подходы будут важны для продвижения в этой области.
Ключевые слова: Arabidopsis thaliana , амилопектин, амилоза, фосфорилирование белков, образование белковых комплексов
Введение
Крахмал представляет собой нерастворимый неструктурный углевод, состоящий из полимеров α-глюкозы. Он синтезируется растениями и водорослями для хранения энергии в плотной, осмотически инертной форме. Крахмал имеет большое значение для человека: он служит основным источником углеводов в сбалансированной диете и возобновляемым сырьем для промышленности.Например, крахмал широко используется в качестве загустителя и текстурирующего агента в обработанных пищевых продуктах, поскольку он желатинизируется с образованием паст при нагревании в воде. Крахмальные пасты также находят бесчисленное количество применений в непищевом секторе, например, в производстве бумаги и картона [1, 2], биоразлагаемых пластиков и упаковочных материалов [3], среди прочего.
В зависимости от его биологических функций крахмал часто подразделяется на два типа: переходный крахмал и запасной крахмал. Крахмал, который синтезируется в листьях непосредственно из фотосинтатов в течение дня, обычно определяется как временный крахмал, поскольку он разлагается следующей ночью для поддержания метаболизма, выработки энергии и биосинтеза в отсутствие фотосинтеза.Если это ночное поступление углеводов сокращается — например, у мутантов с нарушенным синтезом крахмала — растения растут медленнее и испытывают острый голод [4]. Крахмал в нефотосинтезирующих тканях, таких как семена, стебли, корни или клубни, обычно хранится в течение более длительных периодов времени и рассматривается как запасной крахмал. Ремобилизация происходит во время прорастания, прорастания или возобновления роста, опять же, когда фотосинтез не может удовлетворить потребность в энергии и углеродном скелете для биосинтеза. Также мутанты с нарушением биосинтеза запасного крахмала часто оказываются в невыгодном положении, а мутантные семена с низким содержанием крахмала или без него могут даже быть нежелательными [5, 6].Именно этот запасной крахмал мы потребляем в пищу и в качестве экстракта для промышленного использования — на него может приходиться 70–80% сухого веса зерен пшеницы и корней маниоки [7, 8].
Крахмалы из разных растительных источников различаются по своим функциональным свойствам (например, температура начала желатинизации, конечная вязкость пасты, образование двухфазных паст или липкость пасты) и, следовательно, по их конечному использованию. Это изменение происходит из-за различий в структуре крахмала, такой как размер гранул крахмала, их состав и молекулярная архитектура составляющих полимеров [9].Тем не менее, экстрагированный крахмал часто необходимо модифицировать с использованием дорогостоящих и иногда вызывающих отходы химических, физических или ферментативных обработок для придания или улучшения требуемых функциональных свойств [10]. Структура крахмала также влияет на его усвояемость в кишечнике. Продукты с пониженной усвояемостью (резистентный крахмал), такие как крахмалы с высоким содержанием амилозы, все больше ценятся из-за их полезного для здоровья эффекта, потенциально служащего профилактической мерой против таких состояний, как колоректальный рак и диабет [11].Понимание биосинтеза крахмала и его взаимосвязи со структурой и функциональностью представляет огромный интерес, поскольку представляет собой предпосылку для целенаправленного улучшения крахмальных культур.
В этом обзоре основное внимание уделяется механизмам биосинтеза крахмала и делается попытка предоставить широкий обзор наших текущих знаний, в то же время подчеркивая недавние достижения. Значительные шаги в наших базовых знаниях были сделаны путем анализа модельных систем, таких как растение Arabidopsis thaliana и одноклеточная зеленая водоросль Chlamydomonas reinhardtii. Хотя их крахмалы не имеют прямого промышленного значения, многие аспекты биосинтеза крахмала, по-видимому, широко сохраняются в кладе Viridiplantae. Таким образом, открытия, сделанные в этих системах, вероятно, будут иметь большое значение. Тем не менее, всегда важно помнить о клеточном и метаболическом контексте, в котором образуется крахмал. Различия в условиях между тканями и видами могут иметь сильное влияние на количество и структуру крахмала. Такие различия могут объяснить, почему в некоторых случаях разные фенотипы возникают в результате сходных генетических нарушений.В конечном итоге будет важно понимать как основной процесс биосинтеза крахмала, так и тканеспецифические факторы, которые на него влияют.
Структура крахмала
Крахмал состоит из двух полимеров глюкозы — амилопектина и амилозы, которые вместе образуют нерастворимые полукристаллические гранулы крахмала (рис.; См. [12] для более полного обзора). Оба полимера состоят из α-1,4-связанных глюкановых цепей, связанных с α-1,6-точками ветвления, но их структура и биосинтез различны.Амилопектин составляет 75–90% крахмалов дикого типа, имеет степень полимеризации (DP) ~ 10 5 и уровень разветвления 4–5% (т. Е. 4–5% его связей составляют α-1 , 6-точки ветвления) [13]. Амилопектин составляет структурную основу и лежит в основе полукристаллической природы крахмала. Амилоза значительно мельче и слабо разветвлена [13]. Считается, что он заполняет пустоты в полукристаллической матрице, образованной амилопектином, вероятно, делая гранулы крахмала более плотными.
Строение и биосинтез крахмала. a Обзор основного пути биосинтеза крахмала. АДФ-глюкозопирофосфорилаза (AGPase) продуцирует АДФ-глюкозу, субстрат синтаз крахмала (SS). Гранулированная синтаза крахмала (GBSS) синтезирует амилозу, в то время как растворимые SS, ферменты разветвления (BE) и фермент разветвления изоамилазного типа (ISA) совместно синтезируют амилопектин. b Молекулярная структура амилозы и амилопектина (согласно кластерной модели), показывающая характер ветвления и образование вторичных структур. Заполненные соединенные кружки представляют индивидуальные глюкозильные остатки. c Выравнивание двойных спиралей амилопектина высокого порядка. Каждое кольцо роста ( справа ) имеет толщину прибл. 200–400 нм и содержит полукристаллическую и аморфную области. Полукристаллическая область состоит из чередующихся кристаллических пластинок (содержащих линейные части цепей) и аморфных пластин (содержащих большинство точек ветвления), которые складываются с периодичностью ~ 9–10.5 нм ( средний ). В зависимости от точной архитектуры амилопектина, дающего начало кластерам, двойные спирали либо образуют плотно упакованный полиморф A-типа, либо менее плотный гексагональный полиморф B-типа ( вверху ). Смесь A и B также возможна и названа полиморфом C-типа (не показан). Рисунок, составленный с использованием частей из [30] (с разрешения Elsevier) и [284] (thearabidopsisbook.org; Авторское право Американского общества биологов растений)
Относительно простая химическая природа крахмала резко контрастирует со структурной сложностью последнего. гранулы крахмала.Эта химическая простота затрудняет получение окончательной структурной информации о глюканах. Вместо этого методы, используемые для определения количества структурных единиц (например, количества точек ветвления или длины цепи), как правило, предоставляют средние показатели, которые маскируют структурную неоднородность. Такие ограничения означают, что мы полагаемся на структурные модели крахмалов. Это неизбежно может повлиять на интерпретацию данных, таких как предпочтения ферментов в отношении субстратов или мутантных фенотипов, и влияет на нашу способность однозначно приписывать определенные функции ферментов.Тем не менее, для многих ферментов данные о различных типах помогли прийти к единому мнению об их роли в синтезе крахмала.
Молекулярная структура амилопектина
Обычно считается, что частота и характер ветвления амилопектина не случайны. Внутри каждой молекулы составляющие цепи классифицируются в соответствии с их связью с другими цепями: A-цепи — это внешние цепи, которые сами по себе не имеют ответвлений, а B-цепи — это те, которые несут одну или несколько ветвей.C-цепь представляет собой одну B-цепь на молекулу, имеющую свободный восстанавливающий конец. В моделях амилопектина точки ветвления сконцентрированы в определенных областях, из которых сегменты линейных цепей отходят, образуя кластеры (рис.). Частотное распределение длин цепей (распределения длин цепей или CLD), выведенное из анализа разветвленных крахмалов, показывает, что большинство цепей содержат от 10 до 20 единиц глюкозы. Они считаются A- и B 1 -цепями (B-цепями, которые участвуют в образовании одного кластера).Однако существуют и более длинные цепочки, которые, как считается, образуют связи между различными кластерами. Обычно считается, что они ориентированы в той же ориентации, что и заполняющие кластеры A- и B 1 -цепи (кластерная модель [14, 15]) и обозначаются как B 2 -, B 3 -, B 4 -цепи для цепочек, охватывающих два, три или четыре кластера соответственно; [16]. Однако они могут быть также ориентированы перпендикулярно кластерам (каркасная модель [17]).
Рентгеновский анализ и электронно-микроскопический анализ показывают, что кластеры складываются с периодичностью ~ 9–10 нм [18, 19].Картины дифракции рентгеновских лучей также показывают, что соседние линейные сегменты цепи внутри кластеров образуют параллельные двойные спирали, причем каждый полный виток имеет 6 единиц глюкозы на цепь и период 2,1 нм. Двойные спирали выстраиваются в плотный полиморф A-типа или менее плотный (и более гидратированный) полиморф B-типа [20, 21] (рис. C). Также наблюдаются крахмалы, содержащие смеси полиморфов A- и B-типа, которые называются полиморфами C-типа. Полиморфы A-типа типичны для зерен злаков и полиморфы B-типа клубневых крахмалов.Однако факторы, ответственные за определение типа полиморфа, полностью не изучены.
Структуры крахмала высокого порядка
Различные микроскопические анализы предполагают уровни организации за пределами 9-нм-повтора (рис. C). Некоторые из самых ранних рисунков и легких микрофотографий гранул крахмала показали концентрические слои внутри гранул. Они были названы «годичными кольцами» из-за внешнего сходства с годичными кольцами деревьев. Обработка гранул растрескавшегося крахмала α-амилазой или кислотой, которая удаляет менее кристаллические области, и анализ с помощью сканирующей электронной микроскопии четко показывают кольца роста как повторяющуюся слоистую структуру с периодом в несколько сотен нанометров.Считается, что каждый из этих устойчивых слоев состоит из множества повторов длиной 9 нм. Предполагается, что восприимчивая аморфная область имеет более низкий порядок [22]. Помимо структуры кольца роста, в полукристаллических областях крахмалов наблюдались сферические блоклеты диаметром от 20 до 500 нм [23]. Они могут представлять собой левостороннюю суперспираль амилопектина, предложенную Остергетелем и ван Брюггеном [24] на основе электронно-оптической томографии и криоэлектронографического анализа.В то время как некоторые структурные особенности крахмала широко распространены, такие как образование и упаковка двойных спиралей и наличие годичных колец, другие остаются менее изученными. Следует помнить о возможном появлении артефактов во время пробоподготовки для многих применяемых методов.
Гранулы крахмала из разных видов и тканей сильно различаются по размеру и форме: от относительно небольших частиц диаметром 0,5–2 мкм в семенах амаранта и плоских дисков в листьях арабидопсиса до гладких сфер размером до 100 мкм в клубневых корнях [25 , 26].Гранулы содержат небольшое количество белка (обычно 0,1–0,7%), который в основном представляет собой гранулированную синтазу крахмала (GBSS), которая производит амилозу, но также и другие ферменты, синтезирующие амилопектин, такие как другие синтазы крахмала (SS) и ферменты разветвления крахмала. (BEs) [27, 28]. Многие крахмалы также содержат следы липидов и фосфатных групп (ковалентно связанных в положении C6 или C3 глюкозы) [27]. Уровень фосфорилирования зерновых крахмалов чрезвычайно низкий. У крахмала листьев арабидопсиса он составляет около 0.05% (т.е. фосфорилируется примерно одна на 2000 единиц глюкозы), в то время как в клубневых крахмалах она может быть во много раз выше (~ 0,5% в картофеле). Фосфорилирование, по-видимому, ограничивается амилопектином и обогащено аморфными участками [29]. Высокое содержание фосфатов является промышленно значимым признаком, поскольку оно связано с повышенной гидратацией гранул и пониженной кристалличностью, что дает крахмальные пасты с более высокой прозрачностью, вязкостью и стабильностью при замораживании-оттаивании ([30] и ссылки в нем).
Ферменты биосинтеза крахмала
Крахмал синтезируется в пластидах — хлоропластах в листьях или специализированных амилопластах в запасающих крахмал тканях основных сельскохозяйственных культур.Иное дело у красных водорослей и глаукофитов; их так называемый флоридовый крахмал синтезируется в цитозоле путем, который механистически отличается от такового в растениях и зеленых водорослях [31, 32]. Синтез крахмала в растениях включает три основных ферментативных активности: во-первых, SS удлиняют невосстанавливающие концы глюкозных цепей, используя аденозин-5′-дифосфат-глюкозу (АДФ-глюкозу) в качестве донора глюкозила; во-вторых, БЭ образуют ответвления из существующих цепей посредством реакций глюканотрансферазы; и, в-третьих, ферменты разветвления (DBE) снова гидролизуют некоторые из разветвлений (рис.а). Несмотря на то, что они представлены в последовательной форме, важно воспринимать их как одновременный, взаимозависимый процесс. Ферменты биосинтеза крахмала хорошо сохраняются у разных видов растений, что предполагает общее происхождение [33]. Основной механизм биосинтеза крахмала напоминает механизм гликогена, водорастворимого α-1,4- и α-1,6-связанного полимера глюкозы, синтезируемого у многих бактерий, грибов и животных. Однако, как описано ниже, биосинтез крахмала является более сложным с точки зрения дублирования и специализации SS и BE, привлечения дополнительных ферментов (т.е.е., DBE) и другие недавно описанные белки, которые могут способствовать образованию полукристаллических гранул крахмала.
АДФ-глюкозопирофосфорилаза (AGPase) обеспечивает субстрат для биосинтеза крахмала
Синтез крахмала начинается с производства АДФ-глюкозы, субстрата для SS. В фотосинтетически активных хлоропластах листьев образование АДФ-глюкозы напрямую связано с циклом Кальвина-Бенсона через превращение фруктозо-6-фосфата в глюкозо-6-фосфат (Glc-6-P) (катализируемое фосфоглюкозоизомеразой) через глюкозу. -1-фосфат (Glc-1-P) (катализируется фосфоглюкомутазой).Затем AGPase (EC 2.7.7.27) катализирует превращение Glc-1-P и АТФ в АДФ-глюкозу и пирофосфат (PP i ). По этому пути примерно 30–50% фотоассимилятов листьев Arabidopsis распадаются на крахмал [34]. Каждая из вышеупомянутых реакций термодинамически обратима. Однако in vivo продукт последней реакции PP i метаболизируется пластидиальной щелочной пирофосфатазой, гидролизуя его с образованием двух молекул ортофосфата (P i ) [35, 36].Это делает синтез АДФ-глюкозы в хлоропласте практически необратимым. Действительно, мутанты Arabidopsis с дефицитом SSIV (описанные ниже), которые не могут использовать АДФ-глюкозу для синтеза крахмала, оказывают сильное влияние на фотосинтетический метаболизм, что объясняется накоплением АДФ-глюкозы и, как следствие, истощением пула аденилата [37].
Синтез АДФ-глюкозы аналогичен в гетеротрофных тканях, где сахароза импортируется из исходных тканей и метаболизируется с образованием гексозофосфатов в цитозоле.Для биосинтеза крахмала как гексозофосфаты (обычно Glc-6-P, хотя также сообщалось о транспорте Glc-1-P), так и АТФ транспортируются в амилопласт, чтобы служить субстратом для синтеза АДФ-глюкозы [38, 39]. Транспорт гексозофосфата происходит в обмен на P i , в то время как транспорт АТФ происходит в обмен на ADP и P i . В эндосперме злаков этот путь отличается: здесь основная активность AGPase обнаруживается в цитозоле, а АДФ-глюкоза импортируется непосредственно в пластиду через специальный, специфичный для злаков подкласс переносчиков адениновых нуклеотидов [40–43].
Синтез АДФ-глюкозы под действием АГФазы часто рассматривается как «обязательный этап» синтеза крахмала. Имеются убедительные доказательства того, что этот шаг регулируется как на транскрипционном, так и на посттрансляционном уровнях, что подробно рассмотрено в другом месте [44]. Вкратце, AGPase представляет собой гетеротетрамер, состоящий из двух больших регуляторных субъединиц и двух небольших каталитических субъединиц. Во многих случаях было продемонстрировано, что фермент аллостерически активируется 3-фосфоглицератом и ингибируется P и (например.грамм. [45, 46]). Кроме того, фермент чувствителен к окислительно-восстановительной регуляции за счет восстановления межмолекулярного дисульфидного мостика, который образуется между остатками цистеина малой субъединицы [47–49]. Считается, что вместе эти регуляторные свойства обеспечивают производство АДФ-глюкозы и, следовательно, крахмала только при наличии достаточного количества субстратов. Было сделано много попыток стимулировать поток к крахмалу путем экспрессии нерегулируемой AGPase из Escherichia coli или planta (например, [50-57]).Это привело к увеличению содержания крахмала по крайней мере в одном сорте картофеля [50], увеличению общего урожая зерна кукурузы [52, 56] и пшеницы [53] и увеличению биомассы клубневых корней в маниоке [55] (обзор см. В [58]). .
Хотя вышеупомянутый путь продукции АДФ-глюкозы хорошо принят, были предложены другие механизмы продукции АДФ-глюкозы (см. [34] и ссылки в нем). Однако эти альтернативные пути требуют подтверждения.
Доменная структура синтаз крахмала (SS)
SS (АДФ-глюкоза: 1,4-α-d-глюкан 4-α-d-глюкозилтрансферазы; EC 2.4.1.21) принадлежат к семейству гликозилтрансфераз (GT) 5 (CAZy [59]). Они катализируют перенос глюкозильного фрагмента АДФ-глюкозы на невосстанавливающий конец (здесь положение C4) существующей глюкозильной цепи, создавая связь α-1,4 и удлиняя цепь. В биосинтезе крахмала участвуют пять классов SS: четыре растворимы в строме (или частично связаны с гранулами), а один почти полностью связан с гранулами. Растворимые SS (SSI, SSII, SSIII и SSIV) участвуют в синтезе амилопектина, в то время как SS, связанный с гранулами (GBSS), отвечает за синтез амилозы.Существует дополнительный предполагаемый класс SS, названный SSV, который последовательно связан с SSIV, но еще не охарактеризован функционально [60].
SSsсостоят из высококонсервативного С-концевого каталитического домена и вариабельного N-концевого удлинения (рис.). Каталитический домен является консервативным между SS и бактериальными гликогенсинтазами, которые также используют АДФ-глюкозу в качестве субстрата, и содержит как домен GT5, так и домен GT1 (CAZy; [61]). Согласно кристаллической структуре Agrobacterium tumefaciens и E.coli гликогенсинтазы, GBSSI риса и SSI ячменя, каталитический домен принимает складку GT-B, с активным сайтом в щели между двумя доменами GT [62–65]. Связывание АДФ-глюкозы, вероятно, включает один или несколько консервативных мотивов Lys-X-Gly-Gly [66–68] и другие консервативные заряженные / полярные остатки [62, 69–72]. N-концевые расширения классов SS различны. В случае SSIII и SSIV эти удлинения, как было показано, участвуют во взаимодействиях белок-белок, потенциально посредством консервативных мотивов coiled-coil [73–75].N-концевая часть SSIII также содержит три консервативных углеводсвязывающих модуля (CBMs), которые участвуют в связывании субстрата [76, 77].
Доменная структура классов синтаз крахмала (SS). SS из Arabidopsis ( At ) по сравнению с гликогенсинтазами из E. coli ( Ec ) и почкующихся дрожжей ( Sc ). Кукуруза ( Zm ) SSIIIa и SSV включены, поскольку они отличаются по своей структуре от ортологов Arabidopsis. Показаны пластидные транзитные пептиды (N-концевые синие прямоугольники ), внутренние повторы ( серые прямоугольники , RPT), углеводно-связывающие модули семейства 25 ( желтые прямоугольники , CBM), домены спиральной спирали ( зеленые прямоугольники. , C), домены гликозилтрансферазы-5 ( черные ящики, , GT5), домены гликозилтрансферазы-1 ( красные прямоугольники, , GT1) и домен гликозилтрансферазы-3 (, оранжевые ячейки, , GT3).Транзитные пептиды были предсказаны с помощью ChloroP [285], мотивы спиральной спирали с помощью Paircoil2 ([286]; значение p <0,05, минимальная длина 21 аминокислота) и всех других мотивов с помощью SMART. Обратите внимание, что длина домена и аннотация зависят от запрашиваемой базы данных. Например, домен GT3 Sc Gsy2P идентифицирован SMART как домен GT1 и здесь вручную переназначен как GT3 [287]. В то время как Sc Gsy2p является гликозилтрансферазой семейства GT3 и использует UDP-глюкозу в качестве субстрата, все другие показанные синтазы являются гликозилтрансферазами семейства GT5 и используют ADP-глюкозу в качестве субстрата.N-концевые области SSIII, содержащие мотивы спиральной спирали и CBM, высоко консервативны среди различных ортологов, но дополнительно содержат внутренние повторы в некоторых случаях (например, в SSIIIa ячменя и пшеницы). Ортологи SSII часто показывали более слабые предсказания спиральной катушки или их отсутствие. At SSV имеет слабо предсказанный транзитный пептид хлоропласта, но не имеет домена GT1 и его функция неизвестна. SSV из большинства других видов, однако, имеют С-концевое удлинение, включая участок, который был обозначен как предполагаемый GT1-подобный домен [60]. Bar 100 аминокислот (AA)
Дупликации генов привели к появлению нескольких изоформ некоторых ферментов. Кодируемые белки имеют высокую степень сходства последовательностей, но часто по-разному экспрессируются, причем специфические изоформы преобладают в эндосперме или вегетативных тканях [78–84]. Например, для каждого класса SS в зерновых существует несколько изоформ (кроме SSI и SSV). Изоформы «а» SSII и SSIII, по-видимому, являются преобладающими изоформами в эндосперме на основании исследований экспрессии и мутантов (далее мы ссылаемся на эти «а» изоформы SSII и SSIII, если не указано иное).У других видов, включая Arabidopsis и растения, имеющие органы, заполненные накопительным крахмалом (например, картофель), существует только одна изоформа для каждого класса.
Гранулированная синтаза крахмала (GBSS) синтезирует амилозу
Мутанты с пониженной активностью GBSS или без нее, так называемые восковые линии , как правило, производят меньше амилозы или вообще не продуцируют ее, например, в эндоспермах кукурузы [85, 86 ], рис [87, 88], пшеница [89], ячмень [90] и амарант [91], корни маниоки [8], картофель [92, 93], семена гороха [94], листья арабидопсиса [95] и С.reinhardtii [96, 97]. Это говорит о том, что GBSS отвечает за синтез амилозы и что никакая другая синтаза не может заменить ее в этой функции.
Вполне вероятно, что GBSS синтезирует амилозу в гранулированном матриксе, образованном амилопектином. Мониторинг распределения амилозы с течением времени в линиях картофеля, в которых экспрессия GBSS и содержание амилозы были подавлены до низкого уровня, позволили предположить, что амилоза была более очевидной по направлению к центру гранулы крахмала и что это ядро, содержащее амилозу, растет вместе с гранулой [98 ].Важно понимать, что хотя и нерастворимы, гранулы гидратированы, и небольшие молекулы, такие как АДФ-глюкоза, очевидно, могут диффундировать в матрикс и использоваться связанными с гранулами белками [99]. Имеются доказательства in vitro, что GBSS действует скорее как процессивный, чем как распределительный, предпочтительно добавляя единицы глюкозы в одну и ту же цепь вместо переключения между цепями [68, 99]. Активность GBSS также сильно увеличивалась при анализе в концентрациях амилопектина, достаточно высоких для того, чтобы произошла спонтанная кристаллизация глюкана [68].Таким образом, GBSS может синтезировать амилозу, удлиняя отдельные цепи глюкана в среде, окружающей кристаллический или кристаллизующийся амилопектин. Его продукт, вероятно, хорошо защищен от ветвления, что объясняет, почему он в значительной степени линейен. Природа праймера, используемого для синтеза амилозы, полностью не выяснена. Радио-мечение гранул крахмала C. reinhardtii позволило предположить, что GBSS сначала удлиняет цепи амилопектина, а затем высвобождает эти цепи во фракцию амилозы [100].Однако у сосудистых растений это может быть иным, поскольку у арабидопсиса не наблюдалось переноса радиоактивной метки с амилопектина на амилозу [101]. Другим праймером могут быть мальтоолигосахариды, присутствие которых, как было показано, увеличивает активность GBSS и увеличивает его специфичность в отношении синтеза амилозы (в отличие от удлинения амилопектиновых цепей) как in vitro [102], так и in vivo [101].
Хотя GBSS почти полностью связан с гранулами [103], он не содержит каких-либо предполагаемых крахмал-связывающих доменов (рис.). Недавно было показано, что консервативный связывающий крахмал белок, снабженный CBM семейства 48 и длинным мотивом coiled-coil [104], взаимодействует с Arabidopsis GBSS через короткий мотив coiled-coil на GBSS [95]. Это взаимодействие было необходимо как для эффективного связывания GBSS с гранулами, так и для синтеза амилозы.
Данные свидетельствуют о том, что GBSS также способствует синтезу амилопектина. В некоторых исследованиях восковидных мутантов сообщалось, что структура амилопектина слегка изменена [91, 105–108], тогда как в других исследованиях — иногда на тех же видах — она казалась нормальной [109–111].Заметным исключением является C. reinhardtii , где отсутствие GBSS вызвало изменение в структуре амилопектина: C. reinhardtii содержит низкомолекулярную фракцию амилопектина с характеристиками окрашивания йода, промежуточными между характеристиками амилопектина и амилозы, которые были отсутствует у мутантов GBSS [96]. Отличительная функция GBSS C. reinhardtii по сравнению с GBSS сосудистых растений может быть объяснена наличием уникального C-концевого хвоста и многократной более высокой специфической активностью [112].Интересно, что двойной мутант C. reinhardtii , дефицитный по SSIII и GBSS, пострадал в гораздо большей степени, чем каждый отдельный мутант, поскольку он продуцировал лишь небольшие количества в основном водорастворимого глюкана, который был почти лишен цепей с (DP выше 40 [97 , 113]. Также ранние работы по синтезу крахмала из клубней картофеля предоставили доказательства потенциальной роли GBSS в синтезе крахмала. В репрессорных линиях ssIII , в которых GBSS был активирован, амилопектин имел популяцию сверхдлинных цепей и крахмала. морфология гранул была изменена [114].Оба признака, вероятно, являются результатом (повышенной) активности GBSS, поскольку они были отменены, когда активность GBSS подавлялась одновременно с SSIII.
Основной синтез амилопектина: синтазы крахмала от SSI до SSIII
Исходя из мутантных фенотипов, каждый класс SS, по-видимому, играет особую роль во время синтеза амилопектина: проще говоря, SSI и SSII, как полагают, образуют короткие цепочки, заполняющие один кластер (т.е. A- и B 1 -цепи), в то время как SSIII предлагается для синтеза более длинных цепей B, охватывающих кластер [115].Напротив, SSIV, по-видимому, меньше участвует в определении структуры амилопектина, но выполняет функцию инициации крахмальных гранул и контроля морфологии гранул [116–118]. Однако в действительности ситуация более сложная: есть случаи функционального перекрытия между ферментами [119], нарушения биосинтеза ферментами, расщепляющими крахмал [120–122], и образования комплексов между ферментами [73, 74, 123–127] . Кроме того, существуют значительные пробелы в нашем фундаментальном понимании того, как каждый класс выполняет свою предполагаемую роль на молекулярном уровне, как описано в следующих разделах.
Относительный вклад каждого класса SS варьируется в разных тканях и между видами, что, как полагают, по крайней мере частично объясняет структурные различия между крахмалом из разных источников. В эндосперме кукурузы SSI и SSIII составляют основные очевидные активности растворимых SS [128], тогда как в листьях кукурузы транскрипт SSI не был обнаружен [129]. Напротив, SSII и SSIII являются основными очевидными растворимыми SS в семенах гороха и клубнях картофеля [130–132], тогда как транскрипты SSI картофеля почти исключительно обнаруживаются в листьях [133].В листьях Arabidopsis SSI является основным растворимым SS, о чем судят по оставшейся SS-активности одиночных мутантов ss , за которыми следуют SSIII и SSII [116, 134]. Несмотря на то, что он выражен на разумном уровне, SSIV, по-видимому, вносит лишь небольшой вклад в общую активность SS [116, 135]. Однако важно отметить, что оценки очевидного вклада SS могут быть смещенными; супрессия SSIII, например, часто сопровождается активацией SSI и / или GBSS, что означает, что сравнение активности общей синтазы в присутствии или в отсутствие SSIII не только отражает вклад SSIII.Кроме того, сами тесты SS могут предпочтительно измерять один класс по сравнению с другим, что приводит к ошибочным оценкам их относительной активности.
Крахмалосинтаза I (SSI)
Потеря активности SSI вызывает отчетливые изменения CLD амилопектина, особенно в отношении цепей A и B 1 , составляющих кластеры. Амилопектин из эндосперма мутантов ssI из риса (в сорте japonica , т.е. рис с неактивным SSIIa и протекающей мутацией в GBSS [136]) и супрессорных линий пшеницы [137] имеет более короткие цепи DP 6 и 7, меньшее количество цепочек из DP 8–12 и больше цепочек примерно из DP 18.Аналогичные изменения наблюдались и в крахмале листьев мутантов ssI Arabidopsis [122, 138]. Поразительно, что цепи, которые истощены у мутантов, имеют ту же длину, что и цепи, которые предпочтительно синтезируются с помощью SSI in vitro. Было показано, что SSI из кукурузы, фасоли и риса предпочитает короткие цепи (обычно DP <10) в качестве субстратов [136, 139, 140], а SSI из арабидопсиса более активен в отношении гликогена, чем амилопектина [138]. Эти находки указывают на то, что SSI удлиняет короткие цепи глюкана, происходящие из действия BE (которые в основном представляют собой DP 6), на несколько единиц глюкана (до DP примерно 8-10).Затем эти цепи, вероятно, дополнительно удлиняются SSII и, возможно, другими SS. Однако, поскольку у мутантов ssI большинство цепей от действия BE все еще являются удлиненными, другие SS, по-видимому, лишь частично зависят от действия SSI.
Интересно, что цепи, удлиненные в отсутствие SSI, кажутся более удлиненными, увеличивая долю цепей вокруг DP 18 за счет более коротких цепей. При использовании модифицированных субстратов гликогена было сообщено, что активность усеченного на N-конце SSI кукурузы резко снижается с увеличением длины внешней цепи, в то время как его связывание с субстратом сильно увеличивается [139].Авторы предложили сценарий, в котором эти характеристики заставят SSI придерживаться удлиненных глюкановых цепей в качестве неактивного фермента, тем самым предотвращая удлинение за счет других синтаз. В самом деле, захват SSI внутри гранулы до некоторой степени обычно наблюдается [128, 136, 140, 141]. Неясно, действительно ли степень связывания SSI с гранулами достаточна для блокирования значительной части его короткоцепочечных продуктов. Более того, другие данные свидетельствуют о том, что локализация SSI в гранулах зависит от связывания крахмала его партнера по взаимодействию, SSII (см. Ниже).
Более поздние данные in vitro показали, что рекомбинантный SSI арабидопсиса может фактически синтезировать цепи до DP 15 при инкубации с мальтогептаозой в качестве праймера [142] и, при содействии BE, был способен продуцировать весь спектр длин цепей, обычно присутствующих в один кристаллический слой амилопектина (т.е. цепи A и B 1 [143]). Укороченная на N-конце версия SSI ячменя даже давала цепи DP> 40 при использовании мальтогексозы в качестве праймера [144]. Таким образом, настоящая причина, по которой некоторые короткие цепи, генерируемые SSI, не удлиняются дальше в условиях дикого типа, остается неясной.
Arabidopsis SSI является редокс-чувствительным и требует восстановительных условий для активности [145]. В соответствии с кристаллической структурой SSI ячменя было предложено образование дисульфидного мостика между цистеинами 126 и 506, блокирующего активный центр [65]. Однако одиночная мутация каждого из этих остатков цистеина не делала SSI полностью редокс-нечувствительным, и мутантный белок C506S терял большую часть своей активности.
Крахмалосинтаза II (SSII)
Эффекты дефицита SSII были охарактеризованы для клубней картофеля [133, 146–148], семян гороха [149], эндосперма пшеницы [150], ячменя [151], риса ( я.например, сорт риса japonica [152, 153]) и кукурузы [154], а также в листьях Arabidopsis [119, 122, 134]. Наблюдаемые фенотипы очень похожи и включают отчетливое изменение тонкой структуры амилопектина: наблюдается повышенное количество цепей вокруг DP 8 и сниженное количество цепей вокруг DP 18, то есть сдвиг в сторону более коротких цепей. Кроме того, мутантные крахмалы ssII часто имеют больше амилозы, измененную морфологию гранул и пониженную кристалличность крахмала. У арабидопсиса также сообщалось, что вместе с крахмалом накапливаются небольшие количества растворимого глюкана [122].
Основываясь на изменениях в CLD, кажется, что SSII удлиняет цепи примерно DP 8 (цепи, удлиненные SSI) до длины около DP 18. Эта прямая интерпретация осложняется тем фактом, что, по крайней мере, в зерновых, SSII взаимодействует с SSI и БЭ класса II (см. Раздел «Роль комплексообразования и фосфорилирования ферментов биосинтеза крахмала»). Следовательно, потеря SSII может иметь плейотропные эффекты на эти ферменты, что затрудняет оценку того, какая часть фенотипа напрямую связана с отсутствием активности SSII.Например, умеренное увеличение амилозы в мутантах ssII может быть вызвано измененной активностью BEII (см. Раздел «Специфика классов BE»). Однако вполне вероятно, что изменения тонкой структуры амилопектина вызваны отсутствием активности SSII. Во-первых, когда рекомбинантный SSII риса инкубировали с амилопектином из мутанта ssII риса , он специфически удлинял аберрантно короткие цепи, так что теперь модифицированный глюкан выглядел более похожим на дикого типа [155]. Во-вторых, потеря активности SSI на фоне мутанта ssII вызвала типичные изменения ssI , что указывает на то, что SSI все еще активен [122, 134, 136].В-третьих, изменения в амилопектине CLD аналогичны у двудольных растений, у которых нет доказательств образования SSII-содержащих комплексов.
Крахмалосинтаза III (SSIII)
По сравнению с SSI и SSII функция SSIII менее ясна. Его предполагаемые функции включают синтез длинных В-цепей, удлинение цепей, заполняющих кластер (частично дублирующая функция с SSII) и регулирование других ферментов биосинтеза крахмала. Кроме того, SSIII важен для инициации гранул крахмала, по крайней мере, в отсутствие SSIV.В соответствии с мнением о универсальной роли SSIII проявляется как основная активность растворимых SS во всех растениях и тканях, которые были проанализированы на сегодняшний день. Он также несет самое длинное N-концевое удлинение среди всех SS, которое несет домены связывания крахмала и предсказанные домены coiled-coil (Fig.).
Вероятно, наиболее хорошо охарактеризованная функция SSIII заключается в синтезе длинных, охватывающих кластеры B-цепей (например, B 2 , B 3 и т. Д.). Меньшее количество таких цепей наблюдалось у мутантных крахмалов ssIII из клубней картофеля [148], эндоспермов кукурузы [109, 156] и риса [157], а также у C.reinhardtii [97]. Из охарактеризованных мутантов ssIII только мутант ssIII Arabidopsis [158] и мутант ячменя amo1 не проявляют этого фенотипа (обратите внимание, что мутация amo1 не отменяет активность SSIII [159]). .
Изменения профиля короткой цепи амилопектина от мутантов ssIII [147, 148, 157, 158, 160] указывают на то, что SSIII также участвует в синтезе коротких цепей A и B. Эти изменения незначительны по сравнению с изменениями, вызванными отсутствием SSI или SSII.В отсутствие SSII, однако, дополнительная потеря SSIII значительно усиливает мутантный фенотип в зернах риса [80], клубнях картофеля [147, 148] и листьях Arabidopsis [119], подтверждая частично повторяющиеся функции между этими двумя SS. Двойной мутант ssII / ssIII Arabidopsis продуцировал крошечные количества глюканов с резко укороченными цепями, часть которых была растворима в воде [119, 122]. В клубнях картофеля комбинированная репрессия SSII, SSIII и GBSS привела к получению крахмала без амилозы с короткоцепочечным амилопектином, гели которого не подвергались ретроградации при повторных циклах замораживания-оттаивания — предпочтительная характеристика для пищевой промышленности [161].
Мутанты Arabidopsis ssIII , как сообщалось, обладают повышенной общей активностью растворимого SS [158] и производят больше крахмала в течение дня [117, 158] в листе, что позволяет предположить, что он имеет отрицательную регуляторную функцию в отношении других крахмалов. -биосинтетические ферменты. Однако в других исследованиях сообщалось о незначительном снижении активности растворимых СС [117, 134] и уровней крахмала [122, 134]. Увеличение уровней SSI и / или GBSS наблюдалось в эндоспермах ssIII кукурузы [128] и риса [157] и у C.reinhardtii [97]. Поэтому возможно, что некоторые изменения в тонкой структуре амилопектина вызваны повышенными уровнями SSI. Более того, наблюдаемое увеличение содержания амилозы в крахмале ssIII из кукурузы [156, 162] и риса [111, 157] может быть связано с повышенной активностью GBSS. Повышенное действие GBSS также может быть причиной других фенотипов, связанных с дефицитом SSIII, таких как повышенное количество сверхдлинных амилопектиновых цепей и трещинных гранул [114, 157]; репрессия GBSS в дополнение к SSIII в клубнях картофеля подавляла эти аспекты фенотипа ssIII [114].
Инициирование образования крахмальных гранул: синтаза крахмала IV (SSIV) и другие факторы
На сегодняшний день мутанты с пониженной активностью SSIV описаны только у риса [80, 163] и арабидопсиса [116–118]. У риса есть две изоформы SSIV: Os SSIVa, которая мало экспрессируется в листьях и эндосперме, и Os SSIVb, которая обычно экспрессируется выше [79, 80]. Ни одиночные репрессоры Os SSIVa или Os SSIVb [80], ни нулевые мутанты Os SSIVb (в разновидности japonica ; [163]) не обнаруживают заметных изменений в содержании или структуре крахмала в эндосперме семян.Напротив, нулевые мутанты единственной изоформы SSIV у Arabidopsis демонстрируют изменения дневного содержания крахмала в листьях, имея меньше крахмала в конце дня и больше в конце ночи по сравнению с растениями дикого типа, несмотря на то, что у них нормальный мелкий амилопектин. структура [116]. Мутант ssIV Arabidopsis также имеет значительные изменения в количестве и форме гранул крахмала: вместо примерно шести дискоидных гранул [164] хлоропласты мутантов ssIV имеют ноль, одну или две гранулы, которые увеличены и имеют сферическую форму. менее электронно-плотный центр [116, 118].В молодых листьях ssIV хлоропласты не содержали крахмала [118]. Сверхэкспрессия At SSIV у Arabidopsis увеличивала общее содержание крахмала в листьях, хотя об увеличении количества крахмальных гранул не сообщалось [165].
В целом, Arabidopsis SSIV явно выполняет уникальную функцию в отношении инициации и морфологии гранул крахмала, а также степени накопления крахмала [116, 118]. Интересно, что уровни АДФ-глюкозы (субстрат для SS) у мутантов ssIV увеличены более чем в 50 раз, что позволяет предположить, что ее потребление сильно ограничено [118].Это особенно вероятно в случае хлоропластов без крахмала, где оставшиеся изоформы SS не имеют субстрата глюкана. Напротив, растения Arabidopsis, имеющие SSIV в качестве единственного растворимого SS (т. Е. мутантов ssI / ssII / ssIII ), продуцируют такое же количество гранул на хлоропласт, что и растения дикого типа, несмотря на то, что в целом они содержат лишь небольшое количество аберрантного крахмала [117]. Было бы интересно посмотреть, есть ли аналогичные воздействия на крахмал листа в мутантных и репрессорных линиях риса ssIVb .
Анализ нескольких мутантов SS у Arabidopsis показывает, что SSIII (но не SSI и SSII) требуется для достижения некоторого синтеза крахмала в отсутствие SSIV: мутанты Arabidopsis ssIII / ssIV почти полностью не способны синтезировать крахмал и проявляют хлороз и задержку роста. рост [37, 117].Несмотря на сильно сниженную скорость фотосинтеза, эти мутанты по-прежнему накапливают примерно в 100 раз больше АДФ-глюкозы, нарушая баланс аденилатного пула и, возможно, объясняя наблюдаемые плейотропные дефекты [37]. Последствия потери SSIIIa и SSIVb в рисе japonica были совершенно иными, хотя все же поразительными. Двойной мутант ssIIIa / ssIVb все еще был способен продуцировать значительные количества крахмала эндосперма, но накапливал сферические и рыхлые гранулы вместо обычно наблюдаемых полиэдрических гранул сложного типа [163].Этот фенотип сопровождался неровной поверхностью амилопласта и измененным галактолипидным составом мембран. Изменения в последнем ранее были связаны с переходом от гранул простого к сложному типу у мутанта кукурузы opaque5 [166]. Toyosawa и его коллеги предположили, что SSIVb может быть важным для структурной целостности пока еще не охарактеризованных мембраносодержащих структур, подобных перегородке [163], которые могут формировать плесень внутри амилопластов для придания крахмалу многогранной формы [167].Возможные причины фенотипических различий между рисом ssIIIa / ssIVb и ssIII / ssIV Arabidopsis включают присутствие других изоформ SSIII и SSIV в эндосперме риса и / или механистические различия в хранении и переходном образовании крахмальных гранул.
Функция SSIV и механизмы определения количества и формы гранул остаются загадочными. Биосинтез гликогена у других организмов начинается с самогликозилирования либо гликогенсинтаз (в бактериях; [168]), либо специализированных белков, называемых гликогенинами (в эукариотических клетках; [169, 170]).Введение самопрайминга гликогенсинтазы A. tumefaciens в двойные мутанты Arabidopsis ssIII / ssIV действительно восстановило инициацию множественных крахмальных гранул на хлоропласт, но морфология гранул все еще была такой же, как у ssIV [118]. Попытки подтвердить такую самопрайминговую активность в рекомбинантном At SSIV (и At SSIII) не увенчались успехом [117], хотя было обнаружено, что At SSIII катализирует образование непраймированного глюкана в присутствии АДФ-глюкозы [117].Функция автоглюкозилирования может не потребоваться для объяснения инициации полимеров крахмала в фотоавтотрофных тканях, если идеи о синтезе мальтозы de novo во время фотосинтеза подтвердятся [171, 172]: все четыре растворимых SS из Arabidopsis способны удлинять мальтозу и дольше. мальтоолигосахариды (но не глюкоза) [142].
Arabidopsis SSIV, как сообщается, взаимодействует с двумя белками, ассоциированными с пластоглобулами, фибриллинами FBN1a и FBN1b [75]. Это взаимодействие зависело от присутствия некаталитического N-конца SSIV, предположительно содержащего мотивы спиральной спирали (рис.). Исследования локализации SSIV показывают, что это стромальный белок, слабо нацеленный на отдельные точки внутри хлоропласта — первоначально предполагалось, что это края гранул [117]. Однако совсем недавно сообщалось, что он связан с тилакоидными мембранами [75]. В эндосперме риса SSIVb также имел субпластидную локализацию, преимущественно распределяясь в пространстве между гранулами крахмала, хотя отдельные пятна наблюдались редко [163]. Учитывая его предполагаемую роль в определении количества и морфологии гранул, специфическое расположение SSIV в пластиде вполне может оказаться решающим фактором.В свою очередь, понимание факторов, влияющих на его локализацию — в гранулах или мембранах — может дать совершенно новое понимание контроля образования крахмала.
Дополнительные факторы участвовали в контроле количества гранул крахмала. Ряд белков, которые самоглюкозилируются с использованием UDP-глюкозы, были описаны на протяжении многих лет (например, [173–175]), но ни один из них не имел сигналов нацеливания на пластид, но в основном участвовал в биосинтезе клеточной стенки [176]. О белках, гомологичных гликогенинам, сообщалось у Arabidopsis, и подавление белка, содержащего предсказанный сигнал нацеливания на хлоропласты, по-видимому, уменьшало накопление крахмала [177].Это наблюдение, основанное на окрашивании йодом, не было подтверждено. Интересно, что мутантные фенотипы также предполагают участие α-глюканфосфорилазы и DBE изоамилазы как влияющих на количество гранул крахмала (описано в последующих разделах). Также интересно отметить, что один из белков, участвующих в обратимом фосфорилировании глюкана, избыток крахмала 4 (SEX4), активность которого требуется для разложения крахмала в ночное время, также оказывает сильное влияние на морфологию крахмальных гранул, поскольку его потеря приводит к очень сильному разложению крахмала. крупные круглые гранулы [26].Об этом фенотипе не сообщалось для других мутантов, затронутых деградацией крахмала, и его основа еще не исследована.
Классы и доменная структура ферментов ветвления (BE)
Ферменты разветвления крахмала (EC 2.4.1.18) относятся к суперсемейству ферментов α-амилазы (также называемого семейством гликозидгидролаз 13; CAZy: [59]; недавно рассмотрено [ 178] .Они расщепляют цепь α-1,4-глюкана и переносят отщепленную часть в положение C6 единицы глюкозы из той же или другой цепи, создавая ответвление, связанное с α-1,6.При этом BE генерируют дополнительные субстраты для SS (т. Е. Невосстанавливающие концы цепей). BE имеют общую трехдоменную структуру: N-концевой домен, содержащий CBM семейства 48, центральный каталитический домен α-амилазы, характерный для членов семейства Gh23, и C-концевой домен, присутствующий в некоторых α-амилазах (рис.).
Доменная структура ферментов ветвления (BE). Классы BE можно различать по последовательности, но имеют общую доменную структуру. Изображены пластидные транзитные пептиды (N-концевые синие прямоугольники ), углеводсвязывающие модули семейства 48 ( оранжевые прямоугольники , CBM), каталитические домены семейства α-амилазы ( черные прямоугольники , AMY) и все-β домены, обычно обнаруживаемые на С-конце членов семейства α-амилаз (, синие прямоугольники, , AMY_C).Прогнозы доменов были получены, как показано на рис. Домены coiled-coil не были предсказаны с настройками, описанными на рис. Структура домена сохраняется между классами и ортологами, хотя длина предсказанного домена каталитического семейства α-амилазы варьировалась. Поскольку Arabidopsis не имеет класса I BE, для сравнения включен BEI кукурузы ( Zm BEI), а также BE гликогена из почкующихся дрожжей ( Sc Glc3p) и E. coli ( Ec GlgB). По адресу BE1 является предполагаемым BE класса III, в котором отсутствует CBM48, и еще не было показано, что он демонстрирует активность ветвления. Bar 100 аминокислот (AA)
В соответствии с сходством последовательностей БЭ делятся на ферменты класса I (или семейство B) и класса II (семейство A). Ферменты класса I обычно встречаются в виде единой изоформы у planta, за исключением Arabidopsis, который не содержит BE класса I [179]. Класс II представлен одним геном у картофеля и гороха, но две изоформы — BEIIa и BEIIb — встречаются в злаках. Они имеют различные паттерны экспрессии: в кукурузе, рисе и ячмене экспрессия BEIIb ограничена зерном, тогда как BEIIa обнаруживается во всех тканях, но часто на более низких уровнях [180–183].Arabidopsis также имеет два БЭ типа II (ВЕ2 и ВЕ3), оба из которых экспрессируются в листьях и кажутся в значительной степени функционально избыточными [179].
Гомология белковой последовательности выявила предполагаемый третий класс БЭ. Гены, принадлежащие к этому классу III, были обнаружены у Arabidopsis (BE1; [179]), риса, тополя и кукурузы (BEIII; [78, 184]. Несмотря на высокую гомологию внутри этого класса (около 60%), белки разделяют только около 30% идентичности с БЭ из классов I и II [184]. На сегодняшний день функциональный анализ этого предполагаемого БЭ был проведен только на Arabidopsis, где сначала сообщалось, что нокаут-мутанты имеют фенотип дикого типа [179], а затем должны быть неприкосновенный [185].Ни одна из групп не сообщала об активности ветвления. Необходимы дальнейшие исследования, чтобы выявить каталитическую активность и биологическую функцию этих белков.
Специфические классы ферментов ветвления (BE)
Считается, что конкретное размещение ветвей BEs является основным детерминантом кластерной структуры крахмала (то есть обогащением ветвей в определенных, регулярно расположенных областях). Однако из-за наших ограничений в оценке фактического распределения точек ветвления, исследования специфичности изоформ BE часто больше фокусировались на длине перенесенных цепей, а не на их размещении, в первую очередь путем сравнения CLD глюканов до и после модификации in vitro. пользователя BE.Соответственно, BE класса I имеют тенденцию переносить несколько более длинные цепи, чем BEII, и более активны по отношению к амилозе, тогда как BE класса II обычно предпочитают амилопектин [186–190]. Кроме того, BEI из риса модифицировал амилопектин, ограничивающий фосфорилазу, что позволяет предположить, что этот BEI может переносить уже разветвленные цепи [190, 191]. Получение ограничивающих фосфорилазу CLD после воздействия БЭ позволяет также анализировать модификации внутренних структур ветвления [191]. Это показало, что все рисовые БЭ производят относительно широкое распределение расстояний между новыми и старыми ветвями [191].Напротив, гликоген BE из E. coli ( Ec GlgB), передавая цепи аналогичной длины растительным BEII, отличался тем, что он предпочтительно помещал новые ветви всего в трех единицах глюкозы от существующей точки ветвления [191] . Этот структурный признак также наблюдался, когда Ec, GlgB экспрессировался в линии Arabidopsis, дефицитной по своим эндогенным БЭ (двойные мутанты be2 / be3 ) [192]. Тем не менее, Ec GlgB восстановил синтез нерастворимых глюканов [192, 193], хотя и в меньших количествах и в виде частиц неправильной формы.
Мутанты BEI были проанализированы в эндоспермах кукурузы [194, 195], риса [6, 196] и пшеницы [197], а ген репрессирован в клубнях картофеля [198]. Во всех случаях делеция или уменьшение BEI приводило только к незначительным изменениям в структуре / количестве крахмала в лучшем случае, так что его функция in vivo остается неясной. Снижение большей части или всей активности BEII, однако, вызывало заметные изменения в крахмале и хорошо известном фенотипе амилоза — удлинитель ( ae ). Этот фенотип наблюдался у мутантов beIIb кукурузы [199] и риса [200], у мутантов beIIa твердой пшеницы [201], у мутантов BEII у гороха (давая мутанты с морщинистыми семенами, изученные Грегором Менделем [ 202, 203]) и в линиях двойных репрессоров BEIIa и BEIIb пшеницы и ячменя [204, 205].Фенотип ae характеризуется измененной структурой амилопектина с более длинными внешними и внутренними цепями и, в некоторых случаях, повышенным уровнем амилозы [6, 106, 202, 206–208]. Благодаря своим особым свойствам набухания и желатинизации, а также пониженной перевариваемости в пищеварительном тракте крахмал ae представляет особый интерес как для промышленности, так и для здоровья человека [11, 30]. В зернах риса уровни экспрессии BEIIb коррелировали со структурой амилопектина, а усиление репрессии BEIIb сопровождалось сменой полиморфа A на C на B [207, 209].При сверхэкспрессии BEIIb наблюдались глюканы с более низкой кристалличностью, чем обычно, и также накапливались растворимые глюканы [209].
В клубнях картофеля репрессия BEII привела к амилопектину с более длинными цепями [210], но высокое содержание амилозы наблюдалось только при подавлении генов BE как I, так и II класса [211]. Этот крахмал с высоким содержанием амилозы также имел высокий уровень фосфатов — еще одна особенность, представляющая значительный промышленный интерес. Однако общий выход крахмала сильно снизился [211]. Недавно сообщалось, что сверхэкспрессия BEII в картофеле увеличивает количество коротких амилопектиновых цепей (в основном DP 6), снижает температуру желатинизации крахмала и снижает содержание фосфатов [212].
Некоторые мутации в BE влияют на метаболизм крахмала в листьях. Мутанты кукурузы beIIa имеют нормальный крахмал эндосперма, но в основном заменяют крахмал листа глюканом с низкой молекулярной массой и низкой степенью разветвления [213, 214]. Хотя BEIIa является преобладающим БЭ в листьях кукурузы, транскрипты БЭИ также были обнаружены в листе [214], предположительно отвечающие за остаточный синтез глюкана. У Arabidopsis мутанты, разрушенные в обоих БЭ класса II (ВЕ2 и ВЕ3), демонстрируют полную потерю активности БЭ и не способны продуцировать крахмал.Вместо этого они накапливают большое количество мальтозы [179]. Было высказано предположение, что этот водорастворимый глюкан образовывался во время одновременного синтеза линейных цепей SS и разложения пластидальной α-амилазой 3 (AMY3) и / или β-амилазами.
В совокупности многочисленные исследования BE показывают, что, как и ожидалось, активность ветвления абсолютно необходима для синтеза амилопектина. Они также показывают, что специфические особенности БЭ различаются и что они значительно влияют на окончательную структуру глюкана.Более того, их активность по сравнению с SSs важна, что иллюстрируется изменением структуры глюкана при экспрессии в разной степени [209, 212]. Тем не менее, БЭ не являются единственными детерминантами для синтеза подходящих к кристаллизации глюканов, что продемонстрировано способностью БЭ гликогена E. coli восстанавливать синтез нерастворимых глюканов [192, 193]. Следует также иметь в виду, что БЭ образуют комплексы с другими ферментами биосинтеза крахмала, которые могут влиять на их функции (описанные ниже).
Ферменты разветвления (DBE) и обрезка глюканов
DBE растений гидролизуют α-1,6-связи и высвобождают линейные цепи. Они принадлежат к семейству гликозилгидролаз 13 (CAZy: [59]) и разделяют центральный домен α-амилазы и домен связывания крахмала с BE (рис.,). Далее их можно разделить на два типа: изоамилазы (ISA; E.C. 3.2.1.68) и лимит-декстриназа (LDA; E.C. 3.2.1.41). Эти два типа можно различить по белковым последовательностям и субстратной специфичности, поскольку только LDA могут эффективно разлагать пуллулан [215, 216], дрожжевой глюкан, состоящий из α-1,6-связанных мальтотриозильных единиц (отсюда альтернативное название фермента пуллуланаза или ПУ1).Геномы растений кодируют три класса изоамилазы — ISA1, ISA2 и ISA3 — и один LDA.
Доменная структура ферментов разветвления. Ферменты разветвления Arabidopsis ( At ISA1, At ISA2, At ISA3, At LDA) и фермент разветвления E. coli ( Ec, GlgX) разделяют доменную структуру с BE. Фермент непрямого разветвления из почкующихся дрожжей ( Sc Gdb1p) показан для сравнения. Изображены пластидные транзитные пептиды (N-концевой , синие прямоугольники ), углеводсвязывающие модули семейства 48 ( оранжевые прямоугольники, , CBM), каталитические домены семейства α-амилазы (, черные прямоугольники, , AMY) и домен Pfam. неизвестной функции, DUF3372 ( красный ящик , DUF).Прогнозы доменов были получены, как показано на рис. Домены coiled-coil не были предсказаны с настройками, описанными на рис. Там, где можно было получить полноразмерные последовательности, доменные структуры классов сохранялись среди ортологов у различных видов. Sc Gdb1p содержит три домена фермента, расщепляющего ответвления гликогена (GDE) Pfam (hGDE_N, hGDE_central и GDE_C, от N до C конца). Это характерно для эукариотических GDE, которые объединяют олиго-α-1,4 → α-1,4-глюканотрансферазу (EC 2.4.1.25) и α-1,6-глюкозидаза (EC 3.2.1.33) в рамках одного фермента и обезвоживания гликогена в двухступенчатом механизме. Bar 100 аминокислот (AA)
ISA1 и ISA2 участвуют в разветвлении во время синтеза амилопектина и, таким образом, находятся в центре внимания. Они действуют вместе как гетеромультимерный фермент, и ISA1 дополнительно образует активные гомомультимеры у некоторых видов (описанных ниже) — после этого мы используем «ISA» для обозначения как гомомерной, так и гетеромерной активности. Напротив, ISA3 и LDA главным образом разветвляют крахмал во время его деградации, и мутанты этих ферментов часто имеют крахмал — избыток фенотипов ( пол ) [217–220].Тем не менее, ISA3 и LDA действительно влияют на глюкан, вырабатываемый в отсутствие ISA, хотя они не могут заменить его специфическую функцию [121, 218, 221–223].
Мутанты без ISA частично замещают крахмал водорастворимым полисахаридом в эндоспермах кукурузы [224, 225], риса [226] и ячменя [227], в клубнях картофеля [228], листьях арабидопсиса [120, 218] и в C. reinhardtii [229]. С точки зрения уровня разветвления, длины волны максимального поглощения после окрашивания лица с йодом и молекулярной массы этот водорастворимый глюкан напоминает гликоген и, следовательно, был назван фитогликогеном [230].В большинстве случаев все же производится нерастворимый крахмал, хотя и с небольшими структурными изменениями. Только у C. reinhardtii и japonica риса (т.е. риса с фоном ssII ) мутанты isa1 полностью не способны продуцировать крахмал [226, 229]; во втором случае это зависит от мутировавшего аллеля ssII [231].
Распространенной моделью, объясняющей накопление фитогликогена, является модель «обрезки» [232]. Согласно этой модели, ISA удаляет лишние ответвления из вновь созданной аморфной зоны в «пре-амилопектине», так что только соответствующие цепи удлиняются SS.В отсутствие ISA большое количество разветвлений привело бы к стерическим ограничениям, устранению регулярных структур и синтезу дополнительных ламелей. В соответствии с этим фитогликоген Arabidopsis действительно обогащен ветвями, разделенными небольшими расстояниями [120]. Однако, хотя эта модель рассматривает разветвление с помощью ISA как обязательный этап, тщательное наблюдение за мутантами, не имеющими фермента, позволяет предположить, что это не является строго необходимым для создания компетентного для кристаллизации глюкана: не только большинство мутантов с дефицитом ISA вырабатывают некоторое количество крахмала, часто и то, и другое. крахмал и фитогликоген производятся в одной и той же пластиде, и некоторые типы клеток / ткани оказываются мало затронутыми (например,грамм. пучково-влагалищные клетки из листьев Arabidopsis и кукурузы [120, 233, 234]). Streb et al. показали, что крахмал листьев арабидопсиса полностью замещен фитогликогеном у мутанта isa1 / isa2 / isa3 / lda , лишенного всех DBE [121]. Это предполагает некоторое функциональное перекрытие между DBE — результат, согласующийся с предыдущими наблюдениями на арабидопсисе [218, 223], кукурузе [217] и рисе [221, 222]. Однако дополнительная мутация ферментов, разрушающих крахмал, α-амилазы 3 (AMY3) на фоне мутантного разветвления частично восстанавливает синтез гранул крахмала, показывая, что разветвление, хотя и является частью процесса биосинтеза, не является абсолютным требованием [121].Этот результат согласуется с более ранним наблюдением, что фитогликоген склонен к разрушению ферментов [120], и предполагает, что модификации, производимые деградирующими ферментами, также служат для предотвращения образования нерастворимых гранул крахмала на образующихся молекулах. Недавно генетический анализ арабидопсиса [122] и риса [231] показал, что длина цепей, заполняющих кластер, также влияет на то, образуются ли нерастворимые глюканы в отсутствие ISA; относительное увеличение длинных цепей способствует образованию нерастворимых глюканов, тогда как обогащение более коротких цепей полностью их устраняет.Например, двойные мутанты ssI / isa имеют более длинные цепи A и B 1 и продуцируют значительно больше крахмала, чем одиночные мутанты isa [122] (рис.). Таким образом, кажется, что разветвление облегчает или ускоряет кристаллизацию глюканов, а не формально делает это возможным. Сама кристаллизация в твердых гранулах может, в свою очередь, защитить глюканы от преждевременной деградации амилазами [120, 121].
Модуляция мутантного фенотипа isa синтазами крахмала.Просвечивающие электронные микрофотографии хлоропластов арабидопсиса в конце дня. Растения дикого типа (WT) содержат гранулы крахмала ( черные стрелки, ) в мезофилле и эпидермисе. Мутант isa1 / isa2 накапливает преимущественно фитогликоген ( белая стрелка ) в мезофилле, но все еще производит крахмал в эпидермальных клетках. Дополнительная потеря SSI восстанавливает образование гранул крахмала в мезофилле, скорее всего, из-за относительного увеличения доли более длинных, заполняющих кластеры цепей, которые способствуют кристаллизации, несмотря на аберрантное разветвление.Напротив, дополнительная потеря SSII полностью отменяет синтез нерастворимых глюканов даже в эпидермальных клетках. Это, вероятно, происходит из-за относительного увеличения более коротких цепей в отсутствие SSII, нарушая образование вторичных и третичных структур. Epi эпидермальная клетка, Pal палисадная клетка, Bars 1 мкм. Адаптировано из [122], Авторское право Американского общества биологов растений (http://www.plantphysiol.org)
Интересно, что недавние исследования показывают, что сама ISA может действовать разрушающим образом при некоторых обстоятельствах.Во-первых, экспрессия At ISA1– At ISA2 в E. coli [235] и S. cerevisiae (BP и SCZ, неопубликованные данные) препятствовала накоплению гликогена вместо образования более амилопектин-подобного глюкана. . Во-вторых, гораздо меньше глюканов обнаружено у двойного мутанта ssII / ssIII Arabidopsis по сравнению с тройным мутантом ssII / ssIII / isa [122]. Примечательно, что глюканы, полученные в ssII / ssIII растениях, показали структурное сходство с гликогеном в том, что они имели гораздо более короткие цепи, чем амилопектин дикого типа, и частично были обнаружены в водорастворимой фракции [122].Эти данные показывают, что разветвление под действием ISA для ускорения кристаллизации требует соответствующего глюканового субстрата — преамилопектина с адекватной длиной цепи и / или паттерном разветвления. Если структура глюкана несовместима с (быстрой) кристаллизацией при расщеплении ветвей (например, из-за недостаточно длинных цепей), постоянная доступность может вызвать избыточное расщепление разветвлений и, в конечном итоге, деградацию глюкана.
Чисто разрушающее действие ISA может также объяснить образование множества мелких гранул / частиц у мутантов isa [227, 228, 236].Предполагается, что у Arabidopsis этот фенотип является вторичным эффектом, возникающим в результате усиленной α- и β-амилолитической деградации растворимых глюканов: это может давать небольшие растворимые олигосахариды, которые могут служить праймерами для синтеза глюкана или сайтами зародышеобразования для гранул крахмала [ 121]. Однако небольшие разветвленные олигосахариды, возможно, представляющие собой самую раннюю стадию нормального синтеза глюкана, также могут быть отличными мишенями для разветвления под действием ISA, как упоминалось выше. Удаление ветвей и удаление таких глюканов может ограничить пул потенциальных центров зародышеобразования для гранул крахмала.
Отличительные функции гомо- и гетеромультимеров изоамилазы
В зависимости от вида, ISA1 действует либо исключительно в гетеромультимерных комплексах, образованных между ISA1 и ISA2, либо как гетеромерные, так и гомомерные комплексы. Мутанты Arabidopsis isa1 , isa2 и isa1 / isa2 демонстрируют идентичные фенотипы [120]. Более того, At ISA1 и At ISA2 имеют высокую нативную молекулярную массу, взаимодействуют друг с другом и стабильны только в присутствии своего партнера [120, 235].Каталитическая активность наблюдалась только тогда, когда присутствовали как At, ISA1, так и At ISA2 [235, 237]. Эти данные убедительно свидетельствуют о том, что ISA1 и ISA2 образуют облигатный гетеромультимерный фермент. ISA2 несет в себе замены в аминокислотных остатках, важных для катализа, что, вероятно, делает его некаталитическим. Следовательно, ISA1, вероятно, будет каталитической субъединицей, при этом ISA2 требуется для стабильности фермента и, возможно, для его специфичности и / или регуляторных свойств [235, 238, 239].Точно так же у картофеля был обнаружен только гетеромультимерный комплекс ISA1 и ISA2 [240], хотя ISA1 картофеля был активен сам по себе, когда рекомбинантно экспрессировался как S-tagged белок в E. coli [238].
У других видов очевидно, что ISA1 также действует как гомомультимер. Один или несколько гомомерных комплексов ISA1 были обнаружены в дополнение по крайней мере к одному комплексу ISA1 / ISA2 в зернах риса и кукурузы и в C. reinhardtii [240–242]. В недавней кристаллической структуре C.reinhardtii ISA1 имеет удлиненную структуру и спарен с др. ISA1 хвостом к хвосту, хотя нельзя исключить существование высших мультимеров [242]. Мутанты кукурузы и риса isa2 , которые имеют только гомомер ISA1, обладают сходным синтезом крахмала в эндосперме, как и растения дикого типа, что позволяет предположить, что гомомеров ISA1 достаточно [241, 243]. Однако на фоне мутанта ssIII потеря ISA2 привела к накоплению фитогликогена в зернах кукурузы [160] — интригующее, хотя и необъяснимое наблюдение.Также сообщалось, что гетеромультимер ISA1 / ISA2 является более термостабильным и имеет более высокое сродство к фитогликогену, чем один ISA1 [240]. Таким образом, как на каталитические свойства, так и на стабильность ISA1, по-видимому, влияет ISA2, и как гомомерные, так и гетеромерные ISA могут потребоваться для оптимального синтеза амилопектина, по крайней мере, в некоторых условиях.
Роль комплексообразования и фосфорилирования ферментов биосинтеза крахмала
За последнее десятилетие появилось все больше свидетельств образования комплексов между ферментами биосинтеза крахмала и фосфорилирования этих ферментов.Недавно были идентифицированы два дополнительных комплекса, которые отличаются тем, что они включают белки без очевидных каталитических функций (см. Раздел «Связывающие крахмал белки и другие новые белки»). В раннем исследовании, посвященном этой теме, коиммунопреципитация показала, что BEIIb взаимодействует с BEI и фосфорилазой крахмала в эндосперме пшеницы [123]. Обработка белкового экстракта щелочной фосфатазой устраняла эти взаимодействия, тогда как инкубация с АТФ имела противоположный эффект.Таким образом, предполагается, что фосфорилирование является предпосылкой для образования комплекса, что согласуется с наблюдением, что все БЭ пшеницы могут фосфорилироваться [123]. Позже эксклюзионная хроматография (SEC) экстрактов амилопластов эндосперма кукурузы показала, что SSII, SSIII, BEIIa, BEIIb встречаются в различной степени как немономерные формы. Тесты на парное взаимодействие с помощью иммунопреципитации, аффинной очистки и двугибридного дрожжевого теста позволили предположить, что фосфорилированный комплекс SSIII, SSII, BEIIa, BEIIb (и, возможно, SSI) и второй аналогичный комплекс без SSIII [73, 74].Между тем, Tetlow с соавторами сообщили о других зависимых от фосфорилирования комплексах в эндосперме пшеницы, в том числе о комплексе между SSI, SSII и BEIIa или BEIIb [124]. Это наблюдение было также основано на попарной коиммунопреципитации, аналогичных профилях элюирования, полученных с использованием SEC, и последующем вестерн-блоттинге поперечно-сшитых фракций.
Ассоциация SSI, SSII и БЭ класса II, по-видимому, сохраняется, по крайней мере, в эндосперме злаков, что подтверждается также у кукурузы [125], ячменя [126] и риса [127].В семенах пшеницы и ячменя, где BEIIa и BEIIb выполняют повторяющиеся функции, этот комплекс содержит любой из них, а в ячмене, возможно, оба [124, 126]. В эндосперме кукурузы и риса, где BEIIb является доминирующим BE, BEIIa обычно не связан с комплексом [125, 127].
Знание о существовании этих белковых комплексов неизбежно поднимает вопрос об их значении. В частности, важно понимать, чем функции ферментов в комплексе различаются по сравнению с функциями их мономерных форм.Эта информация сильно повлияет на интерпретацию мутантных фенотипов и, следовательно, на наше понимание роли отдельных ферментов: если отсутствие одного фермента отменяет образование всего комплекса и изменяет активность оставшихся теперь исключительно мономерных или частично комплексных ферментов, результирующий фенотип будет результатом не только отсутствия активности фермента, но также и изменения биосинтетического аппарата в целом.
В мутантах ssII SSI и взаимодействующий (е) BEII (ы) постоянно появляются меньше во фракции, связанной с гранулами [150, 151, 244].Это также наблюдалось в связывающем крахмал мутанте SSII [245]. Более того, в отсутствие BEIIb у кукурузы в эндосперме образуется модифицированный комплекс: SSI и SSII теперь взаимодействуют с BEI, BEIIa и фосфорилазой [125, 246]. Точно так же репрессированные BEIIa или BEIIb заменялись на BEI и фосфорилазу в комплексе в ячмене [126]. В обоих случаях замена белка отражалась на профиле белков, связанных с гранулами: новые комплексные ферменты становились связанными с гранулами вместе со своими комплексными партнерами и GBSS [125, 126].Это говорит о том, что белки в комплексе обладают более высоким сродством к крахмалу. Тем не менее, стоит отметить, что и BEIIb, и SSI могут связывать крахмал самостоятельно [139, 245, 246], и остается неясным, требуется ли тесная ассоциация или даже захват для оптимальной активности.
Потенциальная биологическая функция для образования комплекса может заключаться в передаче субстратов от одного фермента к другому — вновь созданная ветвь, возникающая в результате действия BE, может быть непосредственно удлинена сначала SSI, а затем SSII, тем самым повышая общую эффективность процесса.Также возможно, что комплекс придает ферментативную специфичность; например, стерические ограничения всего комплекса могут определять длину цепи, которая передается ВЕ, или где именно размещается новая ветвь. Тетлоу и Эмес [115] далее предположили, что образование комплексов может регулировать активность ферментов аллостерически и / или может защищать растущий глюкан от разложения ферментов. Тем не менее, ни одна из этих гипотез еще не проверена.
Сходным образом роль фосфорилирования BE и некоторых SS [28] остается загадочной.Помимо стимулирования образования комплексов, фосфорилирование также увеличивает активность стромальных BEIIa и BEIIb у пшеницы [123]. Мутация консервативных сайтов фосфорилирования BEIIb кукурузы на серин резко снижает активность рекомбинантного белка in vitro [247]. Поскольку эта активация происходит в отсутствие его партнеров по взаимодействию, это показывает, что фосфорилирование не обязательно увеличивает активность BEIIb за счет образования комплексов. Авторы также представили доказательства участия Ca 2+ -зависимых протеинкиназ в фосфорилировании BEIIb.
Очевидно, что в будущем будет большая потребность в детальном биохимическом и структурном анализе этих белковых комплексов. В частности, создание мутантных белков, которые все еще активны, но не могут взаимодействовать со своими ферментативными партнерами, могло бы помочь в установлении важности комплексов.
Дополнительные факторы с предполагаемым влиянием на синтез крахмала
Обратимое фосфорилирование глюканов
Обратимое фосфорилирование крахмала лучше всего охарактеризовано у Arabidopsis, где оно имеет решающее значение для эффективного разложения.Считается, что фосфорилирование глюкозильных единиц в положениях C6 или C3 (осуществляемое глюканом, водной дикиназой, GWD или фосфоглюканом, водной дикиназой, PWD, соответственно [248, 249]) разрушает и дестабилизирует спиральные структуры, образованные цепями глюкана, делая их доступными для разрушающих ферментов, в частности β-амилаз [250]. Поскольку экзо-действующие β-амилазы не могут разрушать прошлые фосфатные группы, их необходимо гидролизовать одновременно с помощью фосфоглюканфосфатаз SEX4 (гидролизуется по C6 и C3 [251]) и Like-SEX4 2 (LSF2; гидролизуется по C3 [252], недавно рассмотрено в [253]).
Однако фосфорилирование глюкана не ограничивается темным периодом. Крахмал из листьев арабидопсиса, полученный в течение одного фотопериода, фосфорилируется по C6 и C3, что свидетельствует о том, что и GWD, и PWD активны в течение дня. Более того, мутанты lsf2 имеют повышенные уровни фосфорилирования C3 в конце дня, хотя эти мутанты почти полностью разрушают свой крахмал в ночное время. Это говорит о том, что фосфатные группы, введенные в течение дня, обычно также удаляются LSF2 [252].Кроме того, было показано, что количество C6-связанного фосфата в крахмале Arabidopsis в конце дня непостоянно и положительно коррелирует с фотопериодом [254]. Инкубация дисков клубней картофеля с глюкозой и радиоактивно меченным ортофосфатом привела к образованию радиоактивно меченного фосфата в крахмале, что также указывает на то, что фосфат включается во время чистого синтеза запасного крахмала [255].
Актуальность фосфорилирования во время синтеза еще не ясна. Крахмалы из мутантов Arabidopsis gwd , которые практически не содержат фосфатов, имеют нормальную внутреннюю структуру, но отличаются своей зернистой поверхностью, которая нерегулярна и имеет более короткие цепи [256].Однако эти различия могут быть вторичными эффектами неполной деградации крахмала в течение ночи и последующего синтеза глюканов поверх этих измененных гранулированных поверхностей. Мутанты Arabidopsis gwd также имеют лишь незначительный чистый синтез крахмала в течение дня. Это может означать, что отсутствие GWD в целом подавляет синтез крахмала [257]. Однако, подавляя GWD в течение продолжительной ночи, Skeffington и его коллеги получили растения без оставшегося крахмала и GWD, и они синтезировали количества крахмала дикого типа на следующий день [257].Таким образом, маловероятно, что GWD требуется непосредственно в процессе синтеза крахмала. Скорее, фосфорилирование глюканов с помощью GWD для эффективной деградации глюкана в течение ночи может иметь важное значение для последующего синтеза крахмала: либо напрямую, обеспечивая надлежащий исходный материал, либо косвенно, влияя на распределение углерода (мутанты gwd голодают в течение ночи, что может влиять на использование фотоассимилируется на следующий день).
α-Глюканфосфорилаза — фосфолиз или синтез?
α-Глюканфосфорилаза (EC 2.4.1.1) катализирует обратимую реакцию (α-1,4-связанная глюкоза) n + глюкозо-1-фосфат ⟷ (α-1,4-связанная глюкоза) n +1 + ортофосфат. У высших растений есть два класса фосфорилазы: пластидиальный и цитозольный. Пластидный класс, названный Pho1, PhoL (из-за его низкого сродства к гликогену) или PHS1, несет вставку из 78 аминокислот рядом с сайтом связывания глюкана и показывает самое высокое сродство к линейным глюканам, особенно мальтоолигосахаридам.Напротив, цитозольный класс, названный Pho2, PhoH или PHS2, предпочитает разветвленные субстраты, такие как гликоген [258, 259].
Функция Pho1 неоднозначна до такой степени, что даже не ясно, действует ли он синтетическим или фосфоролитическим (т. Е. Разлагающим) образом in vivo. Пластидная концентрация ортофосфата обычно превышает концентрацию глюкозо-1-фосфатного многообразия [260], что способствует фосфолизу in vivo. Тем не менее было показано, что рис Pho1 катализирует удлинение цепи, а также ее укорочение при сопоставимых концентрациях in vitro [261, 262].Рисовые мутанты Pho1 продуцируют крахмал эндосперма с немного более короткими цепями, более низкой температурой желатинизации и измененной формой гранул, хотя этот фенотип не был полностью пенетрантным [261]. Дополнительный фенотип наблюдался, когда большинство зерен сморщивалось и содержало мало крахмала, когда растения выращивали при более низких температурах (20 ° C) [261]. Было высказано предположение, что в этих условиях Pho1 может работать в биосинтетическом направлении, помогая инициировать синтез крахмала путем синтеза мальто-олигосахаридных праймеров для SS, что объясняет условную потерю крахмала у мутантов.Однако репрессия Pho1 в ячмене [263] не оказывала очевидного эффекта на инициацию крахмальных гранул, хотя количество гранул не оценивалось.
Тем не менее, экспрессия генов и / или ферментативная активность Pho1 коррелирует с таковой из известных генов биосинтеза крахмала в клубнях картофеля [264] и эндоспермах кукурузы, пшеницы и ячменя [263, 265, 266], что согласуется с ролью в биосинтезе. Действуя в фосфоролитическом направлении, Pho1 может способствовать разложению мальтоолигосахаридов, созданных во время обрезки глюкана изоамилазой, с образованием глюкозо-1-фосфата для последующего обратного превращения в АДФ-глюкозу [267].Учитывая, что Pho1 взаимодействует с BEIIb и BEI в амилопластах пшеницы и кукурузы дикого типа [123, 268] и с SSI, SSIIa и BEI у мутантов beII кукурузы и ячменя [125, 126], Pho1 также может действовать. косвенно, изменяя свою ферментативную активность или непосредственно на их продуктах.
В C. reinhardtii , который отличается от сосудистых растений наличием двух пластидных изоформ, PhoA и PhoB, потеря PhoB привела к снижению содержания крахмала. Оставшийся крахмал, кроме того, имел более высокое относительное содержание амилозы и структурные модификации [267].Тем не менее, этот фенотип был ограничен условиями азотного голодания, когда поток в крахмал наиболее высок. PhoB также больше похож на цитозольный, чем на пластидный класс сосудистых растений, поскольку он лишен типичной вставки последовательности и имеет низкое сродство к мальто-олигосахаридам [267].
Также было предположение, что PhoI может участвовать в разложении крахмала. Однако не было отмечено значительного влияния на содержание крахмала в листьях при репрессии листовой изоформы Pho1 у картофеля [269] или у мутанта pho1 Arabidopsis [270–272].Хотя эти отчеты указывают на существенную роль во время деградации преходящего крахмала, локальное накопление крахмала наблюдалось вокруг поражений листьев, что привело к предположениям о специфической роли фосфорилазы в обеспечении респираторных субстратов во время стрессовых реакций [270, 273]. Совсем недавно генетические доказательства участия Arabidopsis Pho1 в деградации крахмала были получены на основе дефицита Pho1 на фоне мутантов ssIV и мутантов с нарушенным метаболизмом мальтозы [271, 272].
Белки, связывающие крахмал, и другие новые белки
В последние годы было идентифицировано несколько новых белков, которые влияют на структуру и / или количество крахмала. В некоторых случаях воздействие на крахмал может быть косвенным, в то время как в других белки могут быть частью еще не открытого аспекта процесса биосинтеза крахмала. В любом случае анализ лежащих в основе механизмов может дать новое понимание аспектов биосинтеза крахмала, которые выходят за рамки основного ферментного аппарата, таких как регуляция синтеза крахмала или влияние пластидного окружения на образование и морфологию гранул.
Peng и его коллеги описали рисовый Floury Endosperm6 (FLO6), пластидный белок с CBM48 и способностью связывать крахмал [274]. Зерна, дефицитные по FLO6, откладывают различные гранулы крахмала с изменениями морфологии и структуры амилопектина [274]. Интересно, что FLO6 физически взаимодействовал с ISA1, который сам по себе не связывался с крахмалом. Было высказано предположение, что FLO6 обеспечивает связывание крахмала с ISA1 и что фенотип flo6 является результатом измененной функции ISA1.Это важная гипотеза для проверки, учитывая важность, приписываемую ISA в сокращении образующегося амилопектина для облегчения его кристаллизации.
Другой пластидный белок, содержащий CBM48, Protein Targeting To Starch (PTST), был описан в Arabidopsis [95, 104]. В дополнение к CBM, PTST содержит предсказанные мотивы coiled-coil. Было показано, что PTST связывается с GBSS посредством взаимодействия через мотив спиральной спирали на GBSS и что это взаимодействие позволяет GBSS связываться с гранулами крахмала: потеря PTST приводит к потере GBSS из гранул in vivo и потере синтеза амилозы [ 95].В дополнение к биотехнологической значимости (как высокое, так и низкое содержание амилозы являются желанными характеристиками крахмала), FLO6 и PTST, таким образом, представляют собой захватывающие примеры неферментативных белков, которые, по-видимому, играют важную роль в нацеливании или модуляции активности ферментов биосинтеза крахмала.
Другой класс неферментативных белков совсем недавно был обнаружен у арабидопсиса. Обозначенный Early Starvation1 (EST1), этот связывающий пластидный крахмал белок был идентифицирован, потому что у мутантов, у которых он отсутствует, обнаруживается преждевременное истощение переходного крахмала в ночное время [275].Это, по-видимому, происходит из-за изменения морфологии крахмальных гранул, что приводит к несоответствующей деградации крахмала в дневное время и ускоренной неконтролируемой деградации крахмала в ночное время. Напротив, растения, сверхэкспрессирующие EST1, развили фенотип с избытком крахмала, указывающий на ингибированное разложение крахмала. Хотя точная молекулярная функция белка неизвестна, генетические данные свидетельствуют против прямого ингибирования деградирующих ферментов EST1. Предполагается, что EST1 может каким-то образом способствовать правильному выравниванию молекул глюкана в гранулированном матриксе — процесс, который до сих пор считался спонтанным.Интересно, что гомолог EST1, подобный EST1 (LEST), по-видимому, выполняет противоположную роль: в то время как , чтобы мутанты не обладали фенотипом дикого типа, сверхэкспрессия приводит к фенотипу с низким содержанием крахмала, подобному таковому у мутантов est1 [ 275].
Изменения морфологии крахмальных гранул в эндоспермах злаков также были связаны с нарушениями других клеточных процессов, включая стресс ER [276], нарушение транспортировки запасных белков [277] и изменение липидного состава мембран пластид [166].Эти наблюдения подчеркивают зависимость образования гранул крахмала от общей эффективности функционирования пластид и клеток. Локализованное в пластидах некондиционное крахмальное зерно4 (SSG4) также может попадать в эту категорию. Мутанты, лишенные этого белка, имеют увеличенные зерна крахмала и пластиды, потенциально возникающие из-за дисфункции во время развития пластид [278, 279]. Недавно сообщалось, что потеря другого стромального белка, Floury Endosperm7 (FLO7), вызывает структурные и морфологические изменения в крахмале.Этот фенотип был ограничен периферией семян риса, где также преимущественно экспрессируется FLO7, но функция FLO7 остается неизвестной [280].
Наконец, другие исследования начали раскрывать регуляторные факторы, которые, по-видимому, влияют на структуру и / или содержание крахмала, контролируя экспрессию генов биосинтеза крахмала. Например, факторы транскрипции Os bZIP58 и Rice Starch Regulator1 (RSR1), как было показано, регулируют экспрессию ряда генов биосинтеза крахмала в зернах риса, тем самым влияя на структуру крахмала и упаковку гранул [281, 282].Сходным образом Floury Endosperm2 (FLO2), ядерный белок с мотивом TPR, влияет на морфологию крахмальных гранул и экспрессию генов биосинтеза крахмала, предположительно за счет взаимодействия с факторами транскрипции [283].
Этот разнообразный набор недавно идентифицированных белков служит иллюстрацией того, что еще многое предстоит понять о биосинтезе крахмала и клеточном контексте, в котором он происходит. Стоит отметить, что большинство описанных выше белков эволюционно законсервированы у высших растений, и межвидовой анализ будет важным аспектом дальнейших исследований.
Переваривание, всасывание и энергетическая ценность углеводов
Переваривание, всасывание и энергетическая ценность углеводовВведение
Многое было изучено о переваривании и всасывании углеводов за последние 20 лет, и это новое знание во многих отношениях полностью изменило наше представление о пищевых углеводах. Теперь мы знаем, что крахмалы перевариваются не полностью, и действительно, некоторые из них перевариваются довольно плохо.Мы узнали, что неперевариваемые углеводы являются не просто нейтральными наполнителями, но обладают важными физиологическими эффектами и даже вносят энергию в рацион. «Сахар» не вреден для здоровья, и не все крахмалы одинаково влияют на уровень глюкозы и липидов в крови. Однако знания во всех этих областях далеко не полные. Кроме того, до сих пор не решены споры о том, как определять и как измерять пищевые волокна и крахмал, и в разных частях мира используются разные методы.Это представляет собой серьезную проблему для тех, кто несет ответственность за формулирование политики и рекомендаций в отношении пищевых углеводов и того, как определяется энергетическая ценность и углеводный состав пищевых продуктов.
Энергетическая ценность углеводов
Было использовано множество различных методов, чтобы определить, сколько энергии в пищевых продуктах доступно для метаболизма человека, называемой метаболизируемой энергией (ME). Общее количество энергии в пище (TE) может быть определено калориметрическим методом, но ME меньше TE, потому что не вся энергия в пище поглощается, а часть поглощается, но теряется с мочой.Большая часть непоглощенной энергии заканчивается с калом, но некоторая часть теряется с газами и теплом, выделяемыми во время брожения в толстой кишке.
Наиболее распространенным подходом к определению энергетической ценности пищевых продуктов является факторный метод (68), в котором рассчитывается количество энергии, содержащейся в каждом из различных компонентов пищи (например, жир, белок, углевод, алкоголь). и сумма полученных цифр принимается за количество энергии в пище. Определение энергетической ценности углеводов представляет собой концептуальную проблему, потому что углеводы различаются по их валовой энергии на грамм, степени, в которой они перевариваются и усваиваются, а также тому факту, что неперевариваемые углеводы обеспечивают количество энергии, которое зависит от степени, в которой они ферментируются в толстой кишке.Это может варьироваться от 0 до 100%. Были предложены альтернативные эмпирические модели, основанные на уравнениях регрессии, разработанных на основе экспериментов, в которых общее потребление энергии и ее выделение с мочой и стулом измерялись при различных диетах. Здесь метаболическая энергия в рационе равна общему потреблению энергии за вычетом потерь энергии, которые оцениваются по потреблению азота и недоступных углеводов. Утверждалось, что эмпирические модели для определения энергетической ценности рациона более точны, чем факторный подход, поскольку они имеют все меньше и меньше источников ошибок (68).Тем не менее, кажется маловероятным, что факторный подход будет заменен, по крайней мере, в ближайшем будущем, потому что он укоренился в правилах маркировки пищевых продуктов и таблицах пищевых продуктов.
Переваривание и всасывание углеводов
Полисахариды и олигосахариды перед абсорбцией необходимо гидролизовать до составляющих их моносахаридов. Переваривание крахмала начинается с амилазы слюны, но эта активность гораздо менее важна, чем активность амилазы поджелудочной железы в тонком кишечнике.Амилаза гидролизует крахмал, причем основными конечными продуктами являются мальтоза, мальтотриоза и α-декстрины, хотя также образуется некоторое количество глюкозы. Продукты переваривания -амилазы гидролизуются до составляющих их моносахаридов ферментами, экспрессируемыми на границе клеток тонкого кишечника, наиболее важными из которых являются мальтаза, сахароза, изомальтаза и лактаза (69). При типичных изысканных западных диетах переваривание углеводов происходит быстро, а всасывание углеводов происходит в основном в верхних отделах тонкой кишки.Это отражается в наличии ворсинок в виде пальцев в слизистой оболочке верхнего отдела тонкой кишки и более широких и более коротких ворсинок в нижней половине тонкой кишки. Однако переваривание и всасывание углеводов может происходить по всей длине тонкого кишечника и смещается в сторону подвздошной кишки, когда диета содержит менее легко усваиваемые углеводы или когда присутствуют ингибиторы кишечной глюкозидазы, которые могут использоваться для лечения диабета. В этой ситуации верхняя часть тонкой кишки демонстрирует широкие ворсинчатые структуры с листовидными массивами, тогда как в подвздошной кишке ворсинки становятся более длинными и более похожими на пальцы.
Моносахариды
В тонком кишечнике человека активно всасываются только D-глюкоза и D-галактоза. D-фруктоза не всасывается активно, но имеет более высокую скорость диффузии, чем можно было бы ожидать при пассивной диффузии. Натрийзависимый переносчик глюкозы, SGLT1, отвечает за активный транспорт глюкозы или галактозы с эквимолярным количеством натрия против градиента концентрации в цитоплазму энтероцита. Фруктоза поглощается облегченным транспортом глюкозы транспортером 5 (GLUT5).Глюкоза перекачивается из энтероцита во внутриклеточное пространство транспортером глюкозы 2 (GLUT2) (70). Полный механизм всасывания фруктозы в кишечнике человека не изучен. Когда фруктоза вводится отдельно в растворе, у 40-80% субъектов наблюдается мальабсорбция, а некоторые субъекты могут поглощать менее 15 г фруктозы. Метеоризм и диарея являются обычным явлением при приеме внутрь дозы фруктозы более 50 г. Однако, если фруктоза вводится в сочетании с глюкозой или крахмалом, фруктоза полностью всасывается, даже у субъектов, которые плохо усваивают только фруктозу (71).Поскольку фруктоза редко встречается в рационе в отсутствие других углеводов, мальабсорбция фруктозы на самом деле является проблемой только для исследований, связанных с пероральными нагрузками фруктозы.
Дисахариды
Глюкозидазы щеточной каймы кишечника, как правило, индуцируются. Например, есть данные о том, что высокое потребление сахарозы увеличивает постпрандиальный инсулин и желудочный ингибирующий полипептидный ответ на большие дозы пероральной сахарозы (72), что, вероятно, отражает повышенную скорость всасывания из-за индукции кишечной активности сахарозы.Отсутствие глюкозидаз щеточной каймы приводит к неспособности усваивать определенные углеводы. Это происходит редко, за исключением лактазной недостаточности, которая часто встречается у некавказских популяций. Последнее может быть полным или частичным и приводит к снижению способности переваривать и абсорбировать лактозу.
Гликемический индекс
Реакцию глюкозы в крови углеводной пищи можно классифицировать по гликемическому индексу (GI). ГИ считается достоверным показателем биологической ценности пищевых углеводов.Он определяется как гликемический ответ, вызванный 50-граммовой углеводной частью пищи, выраженный как процент от реакции, вызванной 50-граммовой углеводной порцией стандартной пищи (73). Гликемический ответ определяется как увеличивающаяся площадь под кривой ответа на уровень глюкозы в крови без учета площади под концентрацией натощак (то есть площади под кривой) (74-76). Стандартная еда — глюкоза или белый хлеб. Если глюкоза является стандартом (т. Е. ГИ глюкозы = 100), значения ГИ продуктов ниже, чем если бы белый хлеб был стандартом, в 1 раз.38, потому что гликемический ответ глюкозы в 1,38 раза больше, чем у белого хлеба. Были опубликованы значения ГИ для нескольких сотен пищевых продуктов (77,78) (см. Таблицу 8).
Гликемический индекс и смешанное питание
Обоснованность GI была предметом многочисленных споров, в основном из-за предполагаемого неприменимости к смешанным блюдам. Большая часть разногласий возникла из-за применения неподходящих методов для оценки ожидаемого гликемического ответа при смешанном приеме пищи.При правильном применении GI предсказывает с разумной точностью относительные реакции глюкозы в крови для смешанных блюд одного и того же состава, но состоящих из разных углеводных продуктов (79).
Значение гликемического индекса
Есть ряд долгосрочных последствий изменения скорости всасывания, или ГИ, пищевых углеводов. Имеются убедительные доказательства того, что уменьшение ГИ в диете улучшает общий контроль глюкозы в крови у пациентов с диабетом (80) и снижает уровень триглицеридов в сыворотке у пациентов с гипертриглицеридемией (81).
Есть также некоторые свидетельства того, что гликемический индекс имеет отношение к спортивному питанию и регулированию аппетита. Пища с низким ГИ, употребляемая перед длительными физическими упражнениями, увеличивала время выносливости и обеспечивала более высокие концентрации плазменного топлива к концу тренировки (82). Однако продукты с высоким ГИ приводили к более быстрому восполнению мышечного гликогена после упражнений (83).
ТАБЛИЦА 8 Гликемический индекс избранных продуктов (продолжение на следующей странице)
GI * | н ** | GI * | н ** | ||
ВЫПЕЧКА | ЗЕРНА | ||||
Торты | 87 ± 5 | 9 | Ячмень перловый | 36 ± 3 | 4 |
Файлы cookie | 90 ± 3 | 14 | Треснувший ячмень | 72 | 1 |
Сухарики пшеничные | 99 ± 4 | 8 | Гречка | 78 ± 6 | 3 |
Маффины | 88 ± 9 | 8 | Булгур | 68 ± 3 | 4 |
Рисовые лепешки | 123 ± 6 | 2 | Кускус | 93 ± 9 | 2 |
Кукурузная мука | 98 ± 1 | 3 | |||
ХЛЕБ | |||||
Ядро ячменя | 49 ± 5 | 3 | Кукуруза сладкая | 78 ± 2 | 7 |
Ячменная мука | 95 ± 2 | 2 | Просо | 101 | 1 |
Ядро ржи | 71 ± 3 | 6 | Рис, белый | 81 ± 3 | 13 |
Ржаная мука | 92 ± 3 | 10 | Рис с низким содержанием амилозы | 126 ± 4 | 3 |
Хлебцы ржаные | 93 ± 2 | 5 | Рис с высоким содержанием амилозы | 83 ± 5 | 3 |
Белый хлеб | 101 ± 0 | 5 | Рис коричневый | 79 ± 6 | 3 |
Цельнозерновая мука | 99 ± 3 | 12 | Рис растворимый | 128 ± 4 | 2 |
Прочая продукция a | 100 ± 4 | 5 | Рис пропаренный | 68 ± 4 | 13 |
Фирменный рис | 78 ± 2 | 5 | |||
ЗЕРНОВЫЕ ЗАВТРАКИ | Ядра ржи | 48 ± 4 | 3 | ||
Все отруби | 60 ± 7 | 4 | тапиока | 115 ± 9 | 1 |
Кукурузные хлопья | 119 ± 5 | 4 | Кинелса пшеничная | 59 ± 4 | 4 |
Мюсли | 80 ± 14 | 4 | |||
Овсяные отруби | 78 ± 8 | 2 | МОЛОЧНЫЕ ПРОДУКТЫ | ||
Каша овсяная | 87 ± 2 | 8 | Мороженое | 84 ± 9 | 6 |
Воздушный рис | 123 ± 11 | 3 | Молоко цельное | 39 ± 9 | 4 |
Пшеница воздушная | 105 ± 3 | 2 | Молоко обезжиренное | 46 | 1 |
Измельченная пшеница | 99 ± 9 | 3 | Йогурт д | 48 ± 1 | 2 |
Другое, GI³ 80 b | 103 ± 3 | 15 | Йогурт e | 27 ± 7 | 2 |
Другое, GI <80 c | 72 ± 2 | 4 | |||
ФРУКТЫ | БОБЫ | ||||
Яблоко | 52 ± 3 | 4 | Печеные бобы | 69 ± 12 | 2 |
Яблочный сок | 58 ± 1 | 2 | Горох черноглазый | 59 ± 12 | 2 |
Абрикосы сушеные | 44 ± 2 | 2 | Масляная фасоль | 44 ± 3 | 3 |
Абрикосы консервированные | 91 | 1 | нута | 47 ± 2 | 3 |
Банан | 83 ± 6 | 5 | Консервы из нута | 59 ± 1 | 2 |
Банан недозрелый | 51 ± 8 | 2 | Фасоль | 54 ± 8 | 5 |
Банан перезрелый | 82 ± 8 | 2 | фасоль | 42 ± 6 | 7 |
Киви | 75 ± 8 | 2 | Фасоль консервированная | 74 | 1 |
Манго | 80 ± 7 | 2 | Чечевица | 38 ± 3 | 6 |
оранжевый | 62 ± 6 | 4 | Чечевица зеленая | 42 ± 6 | 3 |
Апельсиновый сок | 74 ± 4 | 3 | Чечевица зеленая консервированная | 74 | 1 |
Лапа лапа | 83 ± 3 | 2 | Лимская фасоль | 46 | 1 |
Персик консервированный | 67 ± 12 | 3 | Горошек сушеный зеленый | 56 ± 12 | 2 |
Груша | 54 ± 4 | 4 | Горошек зеленый | 68 ± 7 | 3 |
Другое, GI <80 f | 54 ± 7 | 7 | фасоль пинто | 61 ± 3 | 3 |
Другое, GI³ 80 г | 92 ± 4 | 5 | Соевые бобы | 23 ± 3 | 3 |
Горох колотый, желтый | 45 | 1 | |||
ПАСТА | ЗАКУСКИ | ||||
лингвини | 71 ± 4 | 6 | Желейные бобы | 114 | 1 |
Макароны | 64 | 1 | Спасатели | 100 | 1 |
Макароны в штучной упаковке | 92 | 1 | Шоколад (разный) | 84 ± 14 | 2 |
Спагетти, белые | 59 ± 4 | 10 | Попкорн | 79 | 1 |
Спагетти твердые | 78 ± 7 | 3 | Кукурузные чипсы | 105 ± 2 | 2 |
Спагетти, коричневые | 53 ± 7 | 2 | Картофельные чипсы | 77 ± 4 | 2 |
Другое | 59 ± 3 | 8 | Арахис | 21 ± 12 | 3 |
КАРТОФЕЛЬ | СУПЫ | ||||
Мгновенно | 118 ± 2 | 5 | Фасолевые супы (разные) | 84 ± 7 | 4 |
Запеченный | 121 ± 16 | 4 | Помидор | 54 | 1 |
Новое | 81 ± 8 | 3 | САХАР | ||
Белый, вареный | 80 ± 2 | 3 | Мед | 104 ± 21 | 2 |
Белый, пюре | 100 ± 2 | 3 | Фруктоза | 32 ± 2 | 4 |
Картофель фри | 107 | 1 | Глюкоза | 138 ± 4 | 8 |
сладкий картофель | 77 ± 11 | 2 | Сахароза | 87 ± 2 | 5 |
Ям | 73 | 1 | Лактоза | 65 ± 4 | 2 |
* GI = гликемический индекс (белый хлеб = 100), среднее значение ± SEM средних значений из различных исследований.
** Количество исследований.a Бублик, смесь для начинки, булочка для гамбургера, булочки, тосты мельба.
b Почки отрубей, Отруби Chex, Cheerios, Кукурузные отруби, Corn chex. Пшеничная миска, Crispix, Golden Grahams, Grapenuts, Grapenuts хлопья, Life, Pro stars, Sustain, Team, Total (диапазон GI, 83-127)
c Почки отрубей с псиллиумом, Red River, Special K (Австралия), отруби Sultana (Австралия) (GI диапазон 67-77)
d С сахаром
e Искусственно подслащенный
f Вишня, фруктовый коктейль, грейпфрут, грейпфрутовый сок, виноград, слива, ананасовый сок
г Ананас, изюм, арбуз, султанас, арбуз
Агрономия | Бесплатный полнотекстовый | Понимание структуры крахмала: последние достижения
Амилопектин является основным сильно разветвленным компонентом крахмала.Размер амилопектина намного больше амилозы. Физико-химические методы, такие как светорассеяние, вискозиметрия или ультрацентрифугирование, предполагают, что средневесовая молекулярная масса (M w ) должна быть порядка 10 7 –10 8 Да [163,164,165,166,167], тогда как среднечисленные значения (M n ) всего 10 5 –10 6 Да [7,111,168,169]. Таким образом, индекс полидисперсности (M w / M n ) очень велик [170]. Takeda et al.[171] обнаружили, что амилопектин из разных источников состоит из трех размерных фракций с приблизительным средним DP n — диапазонами 700–2100, 4400–8400 и 13 400–26 500, из которых обычно преобладают большие молекулы.3.2.1. Категории цепей
Амилопектин состоит из множества цепей, которые намного короче основных цепей, обнаруженных в амилозах. Распределение цепей по размерам анализируется после разветвления с помощью SEC [115, 172, 173, 174], высокоэффективной анионообменной хроматографии (HPAEC) [9, 10, 175] или электрофореза углеводов с использованием флуорофора (FACE) [176, 177, 178] (рис. 2a).Цепи амилопектина делятся на две основные группы: короткие (S) и длинные (L) цепи; деление между группами обычно составляет DP ~ 36. Преобладают короткие цепи, а молярное соотношение цепей S: L варьируется в зависимости от крахмала, причем зерновые обычно имеют наибольшее соотношение (таблица 2). Как уже упоминалось, S-цепи образуют двойные спирали, которые участвуют в образовании кристаллов в гранулах крахмала [53]. S-цепи закреплены на L-цепях, которые, таким образом, функционируют как соединяющие цепи и в основном ограничены аморфными ламелями [11].Уже в 1952 г. Peat et al. [179] предложили полезную номенклатуру цепей в амилопектине: A-цепи незамещены, тогда как B-цепи заменены другими цепями, а C-цепь несет единственную восстанавливающую концевую группу макромолекулы, но в остальном подобна B-цепи. Hizukuri [180] обнаружил, что длинные B-цепи (BL) могут быть далее подразделены на B2-цепи (со средним DP 42-48) и B3-цепи (DP примерно 69-75) и в конечном итоге даже более длинные цепи. Основная группа S-цепей состоит из A-цепей и коротких B-цепей (BS), последние названы B1-цепями Хизукури [180].В дополнение к S- и L-цепям, некоторые молекулы амилопектина также содержат сверхдлинные цепи (EL-цепи), иногда называемые сверхдлинными цепями [181]. EL-цепи состоят из нескольких сотен или даже тысяч гликозильных единиц, поэтому они напоминают цепи амилозы и их трудно отличить от амилозы. Они также образуют комплексы с йодом, что приводит к завышению содержания амилозы [182]. EL-цепи были обнаружены у сортов риса индика, но не у сортов японского риса [182], а также в сортах риса, е.г., маниока, картофель, ячмень и пшеница [109,183,184,185,186,187]. Наконец, С-цепь в препаратах амилопектина обычно имеет широкое распределение по размеру между DP 15 ~ 120 с пиком около DP 40 [188]. Поскольку одна молекула амилопектина содержит только одну С-цепь, это показывает, что длина С-цепи сильно варьируется между отдельными молекулами. Внешние цепи определяются как сегменты, простирающиеся от самой внешней ветви цепи до ее невосстанавливающего конца [ 195]. Согласно этому определению, A-цепи являются полностью внешними, тогда как все B-цепи имеют один внешний сегмент, а остальная часть цепи является внутренней.Вся внутренняя цепь от внешнего ответвления до стороны с редуцирующим концом называется общим сегментом внутренней цепи [196], тогда как цепь была определена как сегмент сердцевины цепи, если исключить самый внешний разветвленный остаток [197]. Сегменты цепи между ответвлениями рассматриваются просто как сегменты внутренней цепи [195], и средняя длина этих последних сегментов была единственным внутренним параметром, который можно было измерить до изобретения гель-проникающей хроматографии (таблица 2). внешние цепи с целью изучения структуры внутренней цепи амилопектина.Для этой цели можно использовать два экзо-действующих амилолитических фермента. Фосфорилаза a (фермент в мышцах и печени, участвующий в метаболизме гликогена) последовательно удаляет один остаток глюкозы с невосстанавливающих концов через фосфоролитический механизм, производя глюкозо-1-фосфат [198,199], тогда как β-амилаза продуцирует β-мальтозу путем гидролиза [200]. ] (как упоминалось выше в разделе 3.1.1). Ни один из ферментов не проходит мимо ветвей. Фосфорилаза а продуцирует ϕ-предельный декстрин (ϕ-LD), в котором все A-цепи были восстановлены до мальтотетраозильных шлейфов [201, 202, 203].Если ϕ-LD далее гидролизуется β-амилазой, каждая цепь восстанавливается дополнительным остатком мальтозы, таким образом оставляя A-цепи в виде мальтозильных шлейфов в так называемых ϕ, β-LD [202]. В β-LD длина внешних остатков зависит от того, содержит ли исходный внешний сегмент нечетное или четное количество глюкозильных единиц. Следовательно, A-цепи остаются DP 2 или 3 [204]. Внутренние цепи анализируются после расщепления изоамилазой и / или пуллуланазой (рис. 2b). Следует отметить, что существуют некоторые различия в предпочтении различных субстратов двумя ферментами: пуллуланаза (которая полностью гидролизует полисахарид пуллулан) более эффективно гидролизует остатки мальтозильных цепей, тогда как изоамилаза (которая не обладает активностью против пуллулана) более эффективно гидролизует весь полисахарид пуллулан. амилопектин [205, 206, 207, 208].Глюкозильные ответвления устойчивы к обоим ферментам [209]. Соотношение цепей A: B обычно составляет от 1,0 до 1,4 (таблица 2). Внутренние короткие B-цепи (BS) в профилях внутренних цепей разделяются на две подгруппы: B-цепи «отпечатков пальцев» (B fp ), обладающие типичным профилем «отпечатков пальцев» на хроматограммах в зависимости от растительного источника, и основная группа BS-цепей (BS major ) [9]. Внутренние BL-цепи делятся на те же подгруппы (B2- и B3-цепи), что и единичные цепи в исходном амилопектине.В профиле внутренней цепи они короче, однако, поскольку внешний сегмент был удален, так что разделение между BS- и BL-цепями обычно находится между DP 23 и 28, в зависимости от источника растения (Рисунок 2b) [9]. Также A-цепи были подразделены на две группы. Кратчайшие цепи в профиле единичной цепи всего амилопектина (до удаления внешних цепей, рис. 2a) в DP 6–8 рассматриваются как A-цепи (они, вероятно, слишком короткие, чтобы быть B-цепями), и были названы « fingerprint »A fp -цепи по аналогии с внутренними B fp -цепями из-за их типичных профилей на хроматограммах [203].Остальные A-цепи считались кристаллическими A-цепями (кристалл A ) [189], поскольку было показано, что цепи с DP 210]. Профиль единичной цепи всего амилопектина можно использовать для оценки средняя длина цепи (CL). Комбинируя информацию как из профилей единичной, так и из внутренней цепи, можно также рассчитать среднюю внешнюю, общую внутреннюю и внутреннюю длину цепи (ECL, TICL и ICL, соответственно) (Таблица 2) [202, 203].3.2.3. Разветвленные единицы
Цепи в амилопектине организованы в большие или меньшие группы, и различные группы могут быть выделены с помощью эндодействующих ферментов.Хотя существует множество эндодействующих амилолитических ферментов, лишь немногие из них были использованы для структурного анализа. Поскольку α-амилаза Bacillus amyloliquefaciens является ферментом, который до сих пор в основном использовался для исследования разветвленных структурных частей различных амилопектинов, в этом обзоре рассматриваются только результаты, полученные с помощью этого конкретного фермента. Среди нескольких амилолитических ферментов α-амилаза B. amyloliquefaciens дает наиболее выраженный паттерн эндо-действия [213] и поэтому особенно полезна для выделения более крупных разветвленных α-декстринов из амилопектина.Фермент содержит девять субсайтов, расположенных вокруг каталитического сайта [214, 215]. Реакция эффективна только со всеми субсайтами, заполненными глюкозильными звеньями субстрата. Первоначально скорость реакции очень высока, потому что на этой стадии субстрат содержит несколько длинных сегментов внутренней цепи, которые полностью заполняют субсайты [214]. Однако вскоре этот критерий перестает выполняться, что приводит к довольно резкому снижению скорости реакции. α-Декстрины, выделенные на этой стадии, считаются единичными кластерами цепей [216, 217, 218, 219, 220, 221, 222].Из-за природы субъединиц в ферменте сегменты внутренней цепи между разветвлениями в изолированных кластерах составляют DP 223]. Группы кластеров, названные «доменами», также были выделены путем прерывания гидролиза до того, как закончится быстрая начальная стадия [211, 224, 225, 226]. Поскольку α-амилаза также атакует внешние цепи амилопектина более неконтролируемым образом, оставшиеся внешние сегменты имеют различную длину [227, 228] и удаляются добавлением фосфорилазы и / или β-амилазы.Затем можно структурно охарактеризовать предельные декстрины кластеров или доменов. Распределение кластеров по размерам велико, варьируется от приблизительно DP 15 до DP 500 или более и зависит от источника амилопектина [224, 227, 228, 229]. Наибольшие кластеры обнаружены у амилопектинов 1 и 2 типа, которые включают злаки, средний размер которых составляет около DP 66–73 [229]. Это соответствует среднему количеству цепей (NC) около 11–14. Однако было обнаружено, что у ячменя очень большие грозди с NC 19.5 (таблица 3). В выделенных декстринах количество длинных цепей было уменьшено по сравнению с исходным амилопектином, в кластерах больше, чем в доменах, показывая, что α-амилаза атакует длинные внутренние цепи [224, 226, 227, 228]. В результате образуются новые более короткие цепи, некоторые из которых похожи на уже существующие в амилопектине, другие, по-видимому, представляют новые категории, что особенно важно вокруг характерной бороздки на хроматограммах между короткой и длинной цепями амилопектина.Поэтому цепям в продуктах α-амилолиза были присвоены строчные буквы, чтобы подчеркнуть их отличие от цепей в амилопектине (обозначены заглавными буквами) [229]. Кажущиеся кластеры, выделенные из амилопектинов, очень медленно гидролизуются α-амилазой в конечном итоге до небольших α-LD, которые составляют конечные разветвленные единицы, называемые строительными блоками [224, 230]. Молекулярный состав и детальная структура строительных блоков очень сложны. Тем не менее, концепцию строительных блоков можно рационализировать, чтобы она состояла из различных групп α-LD с увеличивающимся числом цепей.Таким образом, группа 2 состоит из строительных блоков с двумя цепями и имеет DP 5–9, группа 3 состоит из трех цепей (DP 10–14), а группа 4 состоит из четырех цепей (DP 15–19) [231, 232, 233]. Поскольку сложность каждой группы увеличивается с увеличением размера, более крупные строительные блоки были изолированы только в качестве подготовки к курсу. Следовательно, группа 5 состоит из декстринов с 5-7 цепями (в среднем 6 цепей), а группа 6 имеет в среднем ~ 10 цепей и DP> 35 [233]. Цепи в строительных блоках очень короткие: кроме а-цепей (DP 2 и некоторые в DP 3), большинство b-цепей сосредоточено вокруг DP 5–6.В более крупных строительных блоках можно обнаружить следовые количества цепей с DP 12 ~ 15 [233]. Во всех проанализированных на данный момент выборках в кластерах преобладают строительные блоки группы 2: примерно 40–65% всех строительных блоков (на числовой основе) имеют только две цепи. Затем численность уменьшается с увеличением числа цепей, так что группа 6 представляет лишь несколько процентов или меньше блоков, а распределение по размерам удивительно похоже независимо от структурного типа амилопектина (Таблица 3) [191, 211, 223, 229, 230, 231, 232, 234, 235, 236, 237].Следует отметить, что самые большие строительные блоки в группе 6 содержат столько цепочек, сколько обнаружено в большей части кластеров. Однако цепочки в строительных блоках намного короче, а плотность ветвей (DB) выше (14 ~ 20%) [211, 229, 234, 235, 239], чем в кластерах (11 ~ 15%) [211, 225, 229, 230, 234 239] из-за очень коротких расстояний между ветвями. : ICL всего 1,4 ~ 2,3. Большие группы в зерновых обычно состоят из 4-6 строительных блоков, тогда как маленькие группы (много корней и клубней) содержат в среднем 3-5 блоков (Таблица 3) [229].Интересно, что средняя длина цепи между строительными блоками (межблочный CL, IB-CL) последовательно увеличивается в кластерах от амилопектинов типа 1 к типу 4 от IB-CL ~ 5,7 до 8,0 [229]. До сих пор только в нескольких случаях межкластерная CL (IC-CL) была оценена в изолированных доменах из амилопектинов. IC-CL обычно находится между 10–20 [211, 225, 226, 227, 231]. Самая короткая IC-CL (9,5) была оценена у сладкого картофеля [226] и ячменя [231], а самая длинная (IC-CL 27) — у восковидного картофеля [227].При сравнении крахмалов из десяти различных источников было показано, что IB-CL положительно коррелирует с температурой начала желатинизации [240]. Это примечательное открытие предполагает, что межблочные сегменты, и, скорее всего, также межкластерные сегменты, играют ключевую роль, когда дело доходит до вкладов в термические свойства гранул крахмала. строительные блоки, т. е. они состоят из межблочных сегментов, а также сегментов внутриблочной цепочки.B-цепи присваивается номер, который соответствует количеству межблочных сегментов вдоль цепочки (Рисунок 3) [229]. Таким образом, в b0-цепочках отсутствуют межблочные сегменты, поскольку они полностью находятся внутри строительного блока. DP этих цепей составляет 3–6, и, если они не образованы α-амилазой, они соответствуют цепям B fp в исходном амилопектине. b1-цепи содержат один межблочный сегмент. Они имеют DP 7–18 и были подразделены на b1a-цепи (DP 7–10), в которых практически отсутствует внутренний сегмент на стороне восстанавливающего конца цепи, и b1b-цепи (DP 11–18), которые имеют сегмент на стороне восстанавливающего конца, возможно, с шестью глюкозильными звеньями, который проходит через внутренний строительный блок.b2-цепи (DP 19–27) имеют два межблочных сегмента и сегменты, которые проходят через строительные блоки. Наконец, b3-цепи (DP ≥ 28) имеют как минимум три межблочных сегмента [229]. Грубо говоря, можно считать, что каждый строительный блок в b-цепи вносит вклад внутриблоковым сегментом (DP 5 ~ 6) в дополнение к межблоковому сегменту (DP 5 ~ 8), который соединяется со следующим блоком, т. Е. b-цепи теоретически обладают периодичностью около 12 вычетов.3.2.4. Организация структурных единиц в амилопектине
Задача в понимании структуры амилопектина состоит в том, чтобы организовать части информации (в форме цепочек единиц, внутренних цепей и различных разветвленных единиц с их составом цепей) в значимую модель всего макромолекула.Помимо соответствия экспериментальным данным, любая модель должна также соответствовать существующим знаниям о зернистой структуре и, что важно, быть полезной для объяснения свойств крахмала. Более того, любая модель также должна согласовываться с тем, что было достигнуто в области биосинтеза крахмала, или, в конечном итоге, даже улучшать или уточнять нынешнее понимание биосинтеза крахмала (см. Обзор биосинтеза крахмала, сделанный Тетлоу и Эмесом в этом специальном выпуске). На протяжении многих лет было предложено несколько моделей структуры амилопектина, и исторически их развитие следует за развитием новых методов и открытием новых ферментов, полезных для структурных исследований [241].В этом обзоре описываются две модели, а именно «кластерная модель» и «строительная модель базовой сети», первая из которых наиболее широко принята сегодня, но вторая является самой последней моделью, основанной на экспериментальных результатах, ранее недоступных. Модель кластера была первоначально предложена в 1969 г. Никуни [242] и независимо Френчем в 1972 г. [45]. В этой модели короткие цепи в амилопектине образуют кластеры, а длинные цепи соединяют кластеры между собой (рис. 4a). Модель основана в первую очередь на следующих выводах: (i) Кислотная обработка гранул крахмала удаляет в основном аморфные части, тогда как кристаллические части остаются в значительной степени нетронутыми.Анализ распределения молекул по размерам в оставшихся гранулированных остатках (обычно называемых «линзами», если гранулы обрабатывают разбавленной HCl, или «амилодекстринами Нэгели», если их обрабатывают разбавленным H 2 SO 4 ), показывает, что большинство ветвей были удалены, и большая часть декстринов представляет собой линейные цепи с DP 13 ~ 16 [17,47,49,51,243]. Результат предполагает, что ответвления в основном ограничиваются аморфными частями и соединяются с короткими цепями в кристаллических частях.(ii) Периодическая длина 9–10 нм, обнаруженная методом SAXS, соответствует одной аморфной и одной кристаллической ламелле [66], толщина последней близко соответствует DP коротких цепей обработанного кислотой крахмала [69, 142, 143] . (iii) Гель-проникающая хроматография показывает, что амилопектин состоит из двух основных групп цепей (S- и L-цепей) [244]. (iv) Поляризационный крест в гранулах крахмала показывает, что молекулы расположены радиально со всеми цепями, направленными в одном направлении к поверхности гранул [45].Работы Робина и др. [46] и Маннерс и Мэтисон [245] также были в пользу кластерной модели. Кроме того, в качестве этих элементов были предложены множественно разветвленные декстрины, образованные из амилопектина с использованием циклодекстрингликозилтрансферазы из Klebsiella pneumoniae [246], мальтотетраозил-образующего фермента из Pseudomonas stutzeri [247] или α-амилаз из ячменя [248] и Bacillus subtilis [216]. кластеры в амилопектине. Hizukuri [180] обнаружил периодичность в длине цепи 27–28 глюкозильных единиц среди B-цепей амилопектина, что близко соответствует периодической длине 9–10 нм, т.е.е., длина цепи в виде двойной спирали с 6 остатками на виток и шагом 2,1 нм. Поэтому Хизукури [180] предположил, что B2-цепи охватывают два кластера, а B3-цепи охватывают три кластера и т. Д. Однако более поздние результаты поставили под сомнение точность кластерной модели. Сравнивая профили единичных цепей ряда амилопектинов, Hanashiro et al. [10] обнаружили периодичность в длине цепи из 12 остатков глюкозы среди S-цепей, но не среди L-цепей. Эта более короткая периодичность среди S-цепей и отсутствие периодичности среди L-цепей не могут быть объяснены как B-цепи, проникающие в последовательные слои кластеров.Напротив, похоже, что периодичность 12 остатков проистекает из взаимосвязи строительных блоков в изолированных α-декстринах, как объяснено выше. О’Салливан и Перес [249], работая с молекулярным моделированием, показали, что сегменты аморфной цепи, участвующие в соединении двойных спиралей в кристаллических ламеллах в параллельном выравнивании (чтобы соответствовать либо A-, либо B-полиморфу), принимают разнообразные и почти разные формы. перпендикулярные направления к спиралям. Следовательно, очень маловероятно, что L-цепи, проникающие в кристаллическую пластину в виде радиально организованных двойных спиралей, также проходят через аморфную пластину в виде спиралей и в том же радиальном направлении.Бертофт и его сотрудники [225, 229, 230, 232] обнаружили, что структура многократно разветвленных декстринов, продуцируемых α-амилазой из B. amyloliquefaciens, не соответствует ожидаемой структуре кластеров, предложенной в кластерной модели. Модель предсказывает, что изолированные кластеры содержат только короткие цепи, потому что длинные цепи должны быть расщеплены, чтобы освободить кластеры от макромолекулы. Однако изолированные декстрины все еще содержат некоторые длинные цепи, даже если количество L-цепей уменьшено по сравнению с исходным амилопектином.В самом деле, отягчающим обстоятельством является то, что чем меньше кажущиеся изолированные кластеры, тем более длинные цепочки они обычно содержат. Кроме того, соотношение S: L цепей амилопектина предсказывает среднее количество цепей (NC) в изолированных кластерах из определенного образца крахмала, но изолированные α-декстрины чаще всего имеют более низкую NC, чем предсказанное теоретическое значение [229]. Только в случае амилопектинов с внутренней структурой типа 4 (B-кристаллические крахмалы) НК изолированных кластеров согласуется с кластерной моделью.Эти обстоятельства привели к необходимости найти модель, которая лучше соответствовала бы экспериментальным данным, и привели к тому, что теперь известно как модель магистрали строительных блоков [250]. В модели магистрали строительных блоков L-цепи (или BL-цепи) в амилопектины связаны друг с другом и вместе образуют более длинный хребет (рис. 4b). Строительные блоки вытянуты вдоль магистрали и образуют единую часть (эти блоки характеризуются как «внутренние») [250]. Строительные блоки всех видов полностью случайным образом распределяются по магистрали [251].У некоторых амилопектинов, особенно со структурой типа 1, некоторые из более длинных BS-цепей, вероятно, также вовлечены в основную цепь [229]. BS-цепи, вероятно, также образуют более короткие ответвления к основной цепи, которые соединяются со строительными блоками за пределами реальной основы («внешние» строительные блоки) [250], но некоторые крахмалы могут также иметь более длинные ответвления, отходящие от основы [229]. Короткие цепи (S-цепи) отходят от строительных блоков в более или менее перпендикулярном направлении. В гранулах крахмала S-цепи образуют двойные спирали кристаллических ламелей, т.е.е., точно так же, как в кластерной модели (рисунок 4). Однако, в отличие от кластерной модели, основная цепь (более или менее) полностью встроена в аморфную пластинку и простирается вдоль нее, а не пересекает стопки пластин в полукристаллическом «кольце роста». Строительные блоки разделены межблочными сегментами (DP 5-8 в зависимости от источника крахмала, таблица 3), и иногда эти сегменты немного длиннее, образуя межкластерные сегменты, которые α-амилаза B. amyloliquefaciens легко атакует вначале. стадии гидролиза [225,232].Однако очевидно, что фактическая функция межкластерных сегментов такая же, как и межблочных сегментов; они соединяют строительные блоки. Следовательно, разветвленные структурные единицы в модели являются строительными блоками, и читатель, возможно, уже заметил, что кластеры в традиционном понимании не существуют в модели основной цепи. Α-декстрины, которые α-амилаза продуцирует на ранних стадиях гидролиза. поэтому являются только кажущимися кластерами, которые формируются из-за преимущественной атаки на межблочные сегменты с DP ≥ 9.Это объясняет, почему структура α-декстринов не соответствует кластерам кластерной модели. Вместо этого базовая модель объясняет тот факт, что некоторые декстрины все еще содержат длинные цепочки, потому что длинные цепочки не участвуют во взаимодействии каких-либо кластеров (поскольку кластеров не существует). Вместо этого длинные цепи участвуют в соединении строительных блоков, и, в зависимости от случайного размещения длинных межблочных сегментов, высвободившиеся декстрины будут содержать переменное количество строительных блоков, межблочных сегментов и длины цепочек, некоторые из которых соответствуют длинные b3-цепи [229].Это также объясняет очень широкое распределение α-декстринов по размерам. Поскольку последовательность типов строительных блоков вдоль магистрали является случайной, длина как внутриблочных, так и межблочных сегментов вдоль магистрали изменяется случайным образом [251], и, следовательно, нет явной периодичности в длине цепи между L-цепочками. . Это объясняет, почему Hanashiro et al. [10] не обнаружили периодичности DP 12 при DP> 36. Однако среди более коротких цепей (b2- и b1-цепи в α-декстринах) периодичность наблюдается, поскольку эти цепи в основном встречаются в виде коротких боковых цепей. к позвоночнику и имеют четко выраженный IB-CL.Модель остова также объясняет тот факт, что соотношение S: L цепей в амилопектине обычно не коррелирует с NC изолированных кажущихся кластеров, потому что L-цепи имеют другую функцию в этой модели по сравнению с кластерной моделью.3.2.5. Возможное влияние структуры скелета на свойства крахмала и биосинтез
Предполагается, что организация молекул амилопектина в слоях ламелей в гранулах крахмала является довольно случайной в том смысле, что скелет может простираться в любом направлении и, более того, он возможно, далек от прямого вытянутого строения.Фактически, скелеты отдельных молекул могут даже пересекаться друг с другом, образуя сложную сеть [252]. Двойные спирали простираются от этой сети до кристаллической ламели. Это значит, что двойные спирали отдельных молекул сближаются и кристаллизуются в A- или B-полиморфную структуру. Выше было отмечено, что размеры нанокристаллов, которые были изолированы при слабом кислотном гидролизе гранул крахмала и стеблей из кристаллических пластинок [67], настолько велики, что более сотни двойных спиралей вносят вклад в отдельные кристаллы. [68].Сеть основных цепей в аморфной ламелле, таким образом, будет способствовать формированию двойных спиралей для кооперативного образования кристаллитов, что объясняет размеры изолированных нанокристаллов. Также интересно отметить, что О’Салливан и Перес [249] показали, что только очень ограниченные комбинации длин внутренней цепи приводят к параллельному выравниванию между двумя соседними двойными спиралями, обнаруженными в одной и той же макромолекуле. Фактически, комбинаций настолько мало, что очень маловероятно, что двойные спирали из одного кластера, как предполагается в кластерной модели, могли внести вклад в какую-либо кристаллическую структуру.Другими словами: кластеры не нужны для образования кристаллических ламелей. Напротив, они, скорее всего, предотвратят свое образование, будучи слишком сильно разветвленными (как их чаще всего изображают). Интересно, что было показано, что ферменты разветвления необходимы для точного образования гранул полукристаллического крахмала. Без активности разветвления во время биосинтеза амилопектин не «урезается», что приводит к слишком плотному разветвлению и образованию водорастворимого фитогликогена [253].В отличие от кластерной модели, базовая модель строительных блоков имеет сравнительно открытую структуру: как отмечалось выше, наиболее распространенный строительный блок, представляющий около 50% всех блоков и около 25% всех ветвей [230], имеет только две цепи и одну ветвь. Обе стороны этой единственной ветви заняты межблочными (или межкластерными) сегментами (IB-CL 5–8 или более). Эта открытая структура может быть результатом «обрезки» активности ферментов разветвления и предполагает гибкую структуру, в которой довольно длинный сегмент может свободно перемещаться и изгибаться, в отличие от сильно разветвленных кластеров в кластерной модели, что приводит к упорядоченные A- или B-кристаллиты.Когда, кроме того, эти гибкие сегменты являются частью сложной сети в аморфных ламелях, легко понять чрезвычайное набухание, которое гранулы крахмала обладают в теплом водном растворе [90, 254, 255, 256, 257, 258]. Когда двойной спиральный порядок в кристаллических ламелях разрушается (исчезает поляризационный крест) из-за поглощения воды гранулами, которое оказывает на кристаллиты все большее давление, ничто не препятствует набуханию гранул. Однако целостность гранул остается неизменной, потому что скелеты молекул амилопектина растягиваются во всех направлениях (сравните с баллоном), и только когда они теряют контакт друг с другом, гранулы распадаются [252].В присутствии амилозы гранулы крахмала более стабильны, чем восковые гранулы [161, 259, 260]. Стабилизирующий эффект, возможно, обусловлен смешиванием амилозы с остовами амилопектина в аморфных ламеллах [203], что снижает их гибкость и замедляет набухание. Амилоза также может пересекать стопки ламелей [69,70,71], действуя как «клей» между ламелями, что имеет аналогичный эффект. Дональд и его коллеги предположили, что амилопектин ведет себя как жидкий кристалл с боковой цепью. [98 261 262 263].Эта модель предполагает, что двойные спирали связаны с аморфной «основой» кластеров посредством гибких спейсерных рычагов. Гибкость распорных рычагов объясняет такие явления, как отжиг и желатинизация. За исключением кластеров, их модель, по сути, напоминает структуру магистральной сети строительных блоков. В последнем случае межблочные сегменты представляют собой гибкие рычаги. Чем длиннее сегменты, тем выше гибкость. Действительно, Вамадеван и др. [240, 264] показали, что и отжиг, и температура начала желатинизации положительно коррелируют с IB-CL.Другим признаком природы остова является взаимодействие β-предельного декстрина амилопектина с йодом, которое дает слабый цвет на низких длинах волн (λ max составляет около 507 ~ 540 нм) [96 265 266]. Поскольку у β-LD отсутствуют внешние цепи, результат показывает, что основная цепь имеет сегменты, достаточно длинные для взаимодействия с йодом, то есть эти сегменты, которые предположительно являются более длинными межблочными сегментами, образуют одиночные спирали и комплексы включения с йодом. Однако точная природа этих сегментов не выяснена.Интересно отметить, что гранулы крахмала также образуют комплексы с йодом, что свидетельствует о том, что молекулярные компоненты в гранулах обладают сегментами, достаточно гибкими для образования спиралей. Гранулы, содержащие амилозу, окрашиваются в темно-синий цвет из-за комплекса с амилозой [159, 267]. Примечательно, что эти обработанные йодом гранулы обладают очень ограниченным набуханием и выщелачиванием амилозы [140, 257, 268]. Однако амилопектин также взаимодействует, о чем свидетельствует тот факт, что гранулы без амилозы окрашиваются в красный цвет [267]. Поскольку большинство коротких цепей участвуют в образовании двойной спирали внутри кристаллитов (и, вероятно, также слишком короткие, чтобы давать заметный цвет), возможно, что во взаимодействии участвует аморфная основная цепь.В самом деле, это еще более примечательно, поскольку скелет вряд ли сможет превратиться в спираль, когда короткие цепочки будут закреплены на своих местах в кристаллах. Следовательно, предполагается, что спираль существует заранее, и йод либо просто попадает в ее центральную полость, либо спираль существует в более протяженной конформации, которая сокращается при связывании йода. Сообщалось о незначительных различиях в дифрактограмме WAXS гранул крахмала, окрашенных йодом [159, 160, 269], что позволяет предположить, что некоторые движения происходят и отражаются в упаковке двойных спиралей в кристаллитах.Спиральная структура основной цепи также открывает новые возможности для структуры двойных спиралей, образованных короткими цепями в амилопектине. До сих пор большинство, если не все, внимание было сосредоточено на двойных спиралях, образованных А- и В-цепью (рис. 5а). Было показано, что эти спирали стабилизируются точкой ветвления между двумя цепями [53]. Однако другие возможные комбинации могут быть между двумя A- или двумя B-цепями [250], если они расположены достаточно близко друг к другу, например, в строительном блоке. Однако, как уже говорилось, около половины строительных блоков состоит только из двух цепей (одной a- и одной b-цепи).Затем во внешних блоках должна быть сформирована хорошо зарекомендовавшая себя спираль A-B [250]. Однако во внутренних блоках b-цепь является частью основы, а a-цепь является одиночной. Если межблочный сегмент между двумя такими строительными блоками является спиралевидным (обратите внимание, что многие IB-CL имеют ~ 6 витков, т. Е. Спиральный виток), две A-цепи будут смежными в пространстве и могут взаимодействовать, образуя двойную спираль. (Рисунок 5b) [230,250]. В этом случае можно было ожидать сравнительно слабой двойной спирали, так как стабилизирующая ветвь отсутствует.Если скелет частично (или полностью) существует в виде пре-спиральной структуры, он также может сохранять эту конформацию в растворе. Фактически, указаний в этом направлении не наблюдается. Было обнаружено, что гелеобразующие свойства в серии из четырех восковидных рисовых крахмалов сильно различаются [265]. В структурном отношении единственными обнаруженными различиями были крошечные различия в длине внутренней цепи, тогда как длина внешней цепи была сходной. Поэтому был сделан вывод, что небольшие различия во внутренней структуре были причиной различных реологических свойств.Возможно, внешние цепи в одной молекуле могут взаимодействовать с одинарными спиральными внутренними сегментами в других молекулах, вероятно, образуя двойные спирали [265]. Очевидно, что модель строительного блока основной цепи бросает вызов нашему взгляду на биосинтез амилопектина — а оттуда всю гранулу крахмала. Одним из важных ферментов синтеза амилопектина является фермент разветвления крахмала (SBE) [270]. Замечательное действие SBE имеет три аспекта: (i) фермент гидролизует (1,4) -связь в одной цепи, (ii) он переносит высвободившуюся невосстанавливающую концевую часть расщепленного сегмента цепи на другую цепь (или, в конечном итоге, той же цепи), и (iii) он присоединяет сегмент цепи к другой цепи, синтезируя (1,6) -связь.Было показано, что SBE в качестве субстрата предпочитает двойные спирали, а не одиночные цепи [271]. В свете того, что основная цепь, возможно, имеет одинарную спиральную структуру, возникает соблазн предположить, что основная цепь синтезируется из двойной спирали «пред-скелетной цепи». Фактически, результат воздействия SBE на двухспиральную подложку обсуждался Боровским и соавт. [271] уже в 1979 г. и представлена на рис. 6. В своей работе они предположили, что реакция переноса после расщепления первой цепи просто включает перенос на другую цепь двойной спирали (расстояние меньше 10.5 Å, что является приблизительным диаметром двойной спирали). Непрерывное действие фермента приводит к сильно разветвленному остову, при этом некоторые ответвления образуют строительные блоки (A- и B fp -цепи) или более короткие ответвления (основные -цепи BS ) от остова, а другие являются ответвлениями в самом остове ( в основном BL-цепи).В крошечном мире крахмальных зерен больше значит лучше
Изображение крахмальных зерен, полученное с помощью поляризованного микроскопа. Фото: Крис Томас, Milton Contact.
Майкл Мозди
Крахмальные зерна — это крошечные структуры, которые большинство растений образуют как продукты фотосинтеза. По сути, крахмальное зерно — это хорошо упакованный кладезь сахарных единиц глюкозы. Многие растения хранят зерна крахмала в подземных органах, таких как корни, луковицы, клубни и корневища, а также в их семенах и стеблях. Сегодня крахмал является наиболее распространенным источником энергии в нашем рационе, если его употреблять в виде картофеля, пшеницы, кукурузы и риса. В доисторические времена это тоже было правдой.(Прочтите «Хорошие новости об углеводах», чтобы более подробно познакомиться с археоботаниками и их изучением древних диет).Зерна крахмала важны для археологов, потому что они необычайно стабильны и эластичны в течение длительных периодов времени. Они сохраняются на древних артефактах и в археологических записях, тогда как другие биологические структуры не могут быть сохранены (пыльца, семена, фрукты и т. Д.).
Лисбет Лудербак, куратор археологии NHMU и доцент кафедры антропологии U of U, опубликовала статью в июне 2016 года после проведения обширной работы по крахмальным зернам со своими коллегами Николь Херцог и Брюсом Павликом.Они исследовали четыре крахмалистых растения с подземными хранилищами, обнаруженные в рационе доисторических людей на американском Западе: картофель «Четыре угла», желтую лавинную лилию, лилию сего и желтую рябину. В статье они описывают, почему требуется большой образец археологических зерен крахмала, чтобы: а) провести уверенную таксономическую идентификацию растения и б) выявить уникальные особенности зерна.
Чтобы понять их выводы, мы должны погрузиться в мир биологии крахмала.
Really Small
«Вы знаете, когда вы режете картофель, на вашем ноже остаются липкие белые остатки? Это миллиарды зерен крахмала », — объясняет Лаудербэк. Хотя мы видим этот белый мазок (и чувствуем липкую текстуру), на самом деле зерна очень маленькие. На самом деле они микроскопические: всего около 20 микрон (мкм) в длину (микрон или микрометр составляет одну миллионную метра — по сравнению с миллиметром, который составляет одну тысячную метра). Для сравнения: эритроцит имеет диаметр 9 микрон, клетка печени — 20 микрон, а клетка кожи — еще больше — 30 микрон.
Основные характеристики
При правильной микроскопии эти маленькие парни выглядят где-то между медиатором и летающей тарелкой. Louderback использует несколько типов микроскопии. Под растровым электронным микроскопом они имеют матовую поверхность, а общий эффект напоминает лунный пейзаж. Однако при использовании микроскопа в проходящем свете можно увидеть еще несколько деталей на крахмальных зернах.
Слева : зерно крахмала видно с помощью сканирующего электронного микроскопа. Справа : видно в просвечивающий световой микроскоп. Фотографии: Лизбет Лоудербэк
Все зерна крахмала имеют хилум, который является точкой, вокруг которой откладываются слои белка. В клубнях, луковицах и корневищах ворота часто расположены не по центру, а кольца роста выходят наружу из ворот. Из-за этого смещения хилума они могут иметь форму «медиатора».
| Поперечное сечение зерна крахмала, показывающее их трехмерную форму.Фото: Эмма Пиллинг и Элисон Смит, впервые опубликовано в книге «Формирование кольца роста в гранулах крахмала клубней картофеля». Эмма Пиллинг и Элисон М. Смит. Физиология растений 2003, том 132, стр. 365–371. |
Годичные кольца показывают, как собираются зерна. Основная единица — продукт фотосинтеза, глюкоза. Глюкоза — это простой сахар, который может быть соединен вместе в два типа цепей крахмала, амилозу и амилопектин. Амилоза представляет собой довольно неразветвленную и линейную цепь, но амилопектин может быть разветвленным и содержать до ста тысяч единиц глюкозы. Цепи амилопектина имеют форму двойной спирали (не путать с самой известной двойной спиралью ДНК). Пластинки (по существу слои) амилозы и амилопектина упакованы вместе в каждом кольце, сложная трехмерная структура которого до сих пор полностью не изучена.Кольца так плотно и равномерно упакованы, что зерно представляет собой почти кристаллическую структуру. Собираетесь в ближайшее время в путешествие? Считайте крахмал лучшим упаковщиком в природе.
Плотность крахмала приводит к одной из наиболее интересных историй совместной эволюции: для того, чтобы люди действительно достигли питательной ценности крахмала, нам необходимо нагреть и распутать это концентрированное ядро из свернутой глюкозы. Итак, только после открытия огня для приготовления пищи люди не смогут по-настоящему извлечь пользу из углеводов.
Вернемся к характеристикам крахмального зерна: одной из основных отличительных характеристик крахмального зерна является наличие структур двойного лучепреломления. Двулучепреломление — это свойство некоторых материалов разделять свет за счет поляризации на два разных луча. Датский ученый впервые заметил это в кристалле кальцита еще в 1669 году. Как ни странно, когда вы включаете фильтр поляризованного света в просвечивающем световом микроскопе, зерна крахмала имеют контрольный набор «рук» и «ног», которые сходятся вместе на холме. .Это называется крестом вымирания, и он имеет вид «X отмечает точку», когда хилум находится по центру, но на мой взгляд он имеет сверхъестественное сходство с хромосомой в некоторых крахмальных зернах со смещенным центром хилы (хотя они полностью не связаны).
Слева : Двулучепреломление в кристалле кальцита, фото: APN MJM / wikipedia. Центр : Двулучепреломление в крахмальных зернах (крест угасания), фото: Лизбет Лоудербэк. Справа : хромосомы человека, фото: Рассел Кенни / flickrПоследняя особенность, важная для исследования Лудербека, называется продольной трещиной.Эту трещину можно представить как треугольный желоб, выходящий наружу из ворот. Неизвестно, какую функцию выполняет трещина и почему она присутствует только в некоторых крахмальных зернах, а не в других.
Выводы Лаудербека
«Продольные трещины становятся определяющей характеристикой некоторых видов», — утверждает Лаудербэк. Оказалось, что продольная трещина совершенно различна у каждого вида растений, участвовавших в ее исследовании. Они могут быть узкими или широкими, разветвленными или неразветвленными.На рисунке ниже показаны четыре растения, участвовавших в ее исследовании, и то, как у них очень разные продольные трещины.
Четыре зерна крахмала, исследованные в работе Лаудербека, каждое из которых демонстрирует отчетливую продольную трещину. Фотографии: Лизбет Лаудербэк
Хотя продольная трещина является полезным признаком идентификации, она не всегда присутствует. Фактически, только 18% крахмальных зерен в исследовании имели трещины. Лаудербэк и ее коллеги заметили, что это было гораздо более распространено в более крупных зернах, однако, когда изучаются только крупные зерна, это происходит в 50-60% случаев.Более крупные зерна также имели более легко распознаваемые кольца роста и форму.
Важное открытие авторов заключается в том, что археологам нужны большие выборки из их справочного и археологического материала, чтобы точно идентифицировать рассматриваемый вид. Более ранние исследования зерна крахмала полагались на форму и размер только нескольких зерен, чтобы сделать идентификацию, но, как показывает эта статья, каждый вид имеет широкий диапазон размеров зерна, а также некоторые вариации по форме.Если у археологов есть большой образец крахмальных зерен, они могут сосредоточиться на более крупных, которые, как правило, помогают им определить, какие виды присутствуют в образце.
Авторы исследования (слева направо): Николь Херцог, Лисбет Лудербэк и Брюс Павлик. © NHMU.
Louderback с радостью использует это для более точной идентификации зерен крахмала на артефактах из других археологических памятников. В настоящее время она работает над статьей, в которой исследует одно из растений в своем исследовании и то, как люди его использовали, и мы с нетерпением ждем возможности ее прочитать.
Исследование Лаудербека было выполнено благодаря щедрому гранту Национального научного фонда и вице-президента Университета Юты по стимулирующему семенному гранту финансирования исследований. Обсуждаемый документ: Louderback, L.A., Herzog, N.M., Pavlik, B.M., 2016. Новый подход к идентификации гранул крахмала из дикорастущих пищевых растений в засушливой западной части Северной Америки. Крахмал — Stärke 68, 1–7.
Майкл Мозди (Michael Mozdy) — писатель по цифровым наукам в Музее естественной истории штата Юта, входящем в состав Университета штата Юта в Солт-Лейк-Сити.Наша миссия — осветить мир природы и место человека в нем. NHMU — это не только выдающиеся экспонаты для публики, но и исследовательский музей. Учить больше.Спросите эксперта: бобовые и устойчивый крахмал | Источник питания
Эксперт: доктор Гай Кросби
Бобовые, в состав которых входят бобы, горох и чечевица, являются недорогим и здоровым источником белка, калия и сложных углеводов, включая пищевые волокна.
- В среднем бобовые содержат около 20-25% белка по весу в пересчете на сухое вещество, что в 2-3 раза больше белка, чем в пшенице и рисе.Однако они, как правило, содержат мало незаменимой аминокислоты метионина, а иногда и триптофана.
- Бобовые также являются очень хорошим источником пищевых волокон, которые важны для поддержания здоровой функции кишечника.
- Содержание общих углеводов, включая сложные углеводы, колеблется от 65 до 72% по весу в пересчете на сухое вещество, из которых 85% состоит из крахмала, а пищевые волокна составляют от 10 до 20% от веса сушеных бобовых ( 1).
Высокий уровень крахмала и пищевых волокон поднимает очень интересный вопрос.Если большая часть углеводов состоит из крахмала, как бобовые могут быть такими хорошими источниками пищевых волокон? Разве не весь крахмал быстро переваривается до глюкозы? Это было преобладающим представлением о крахмале до 1980-х годов, когда два английских исследователя, Ганс Энглист и Джон Каммингс, обнаружили, что не весь крахмал быстро переваривается в глюкозу в тонком кишечнике (2).
- Они обнаружили, что некоторое количество крахмала устойчиво к перевариванию и попадает в толстую кишку, где большая часть его используется в качестве источника пищи здоровыми бактериями, живущими в нашей толстой кишке.Энглист и Каммингс назвали эту ранее неизвестную форму крахмала «резистентным крахмалом» и пришли к выводу, что он действует аналогично пищевому волокну.
- Существенные исследования, проведенные с момента их открытия, подтвердили, что резистентный крахмал (RS) действует так же, как пищевые волокна в пище (3).
Пищевые продукты содержат две основные формы пищевых волокон:
- Одно из них — нерастворимая клетчатка, состоящая из сложных углеводов, таких как целлюлоза, которые нерастворимы в желудочно-кишечной жидкости. Эта форма клетчатки не расщепляется кишечными бактериями, действует как наполнитель и эффективно уменьшает запоры.
- Другая форма клетчатки растворима в желудочно-кишечной жидкости, образуя густую вязкую жидкость, похожую на мед. Растворимая клетчатка легко метаболизируется кишечными бактериями, которые превращают большую ее часть в небольшие молекулы, называемые короткоцепочечными жирными кислотами (SCFA). Клетки, выстилающие толстую кишку, получают около 60-70% своей энергии из SCFA (4).
Хотя резистентный крахмал не увеличивает вязкость желудочно-кишечной жидкости, он является одним из лучших источников SCFA, помогающих поддерживать здоровье клеток толстой кишки.
- Поскольку резистентный крахмал не метаболизируется в тонком кишечнике, он снижает количество глюкозы, попадающей в кровь, тем самым снижая потребность в инсулине, а также снижает калорийность пищи (5).
- Продукты, содержащие значительный уровень резистентного крахмала, повышают чувство насыщения и имеют более низкий гликемический индекс, вызывая меньшее повышение уровня глюкозы в крови, чем продукты с высоким содержанием крахмала, которые содержат очень мало резистентного крахмала, такие как печеный картофель, рис и белый хлеб (6).Ясно, что не все углеводы одинаковы.
Бобовые — один из лучших источников устойчивого крахмала. Сырые сушеные бобовые содержат около 20-30% устойчивого крахмала по весу (7). Это означает, что почти половина крахмала в сырых бобовых не переваривается.
Почему крахмал устойчив к перевариванию? Небольшая его часть физически недоступна для пищеварительных ферментов. Но большинство из них устойчиво из-за химической структуры крахмала. Крахмал состоит из двух молекул, называемых амилозой и амилопектином.
- Амилоза представляет собой линейную цепочку молекул глюкозы, соединенных встык.
- Амилопектин — это гораздо более крупная молекула с многочисленными ответвлениями коротких цепочек молекул глюкозы, связанных с основной цепью, как ветви дерева, растущие из ствола.
Молекулы крахмала, особенно амилоза, образуют кристаллические области, устойчивые к перевариванию ферментами, переваривающими крахмал, в нашем организме (3). По сравнению с другими продуктами с высоким содержанием крахмала, такими как кукуруза, пшеница и рис, крахмал в бобовых очень богат амилозой, составляя до 40% крахмала, что делает их более устойчивыми к перевариванию.
- Важно понимать, что резистентный крахмал — это не отдельная молекулярная структура, такая как глюкоза или холестерин, а концепция, разработанная для объяснения того, почему некоторая часть крахмала не переваривается (8).
- Количество устойчивого крахмала, указанное в пищевых продуктах, в значительной степени зависит от метода, используемого для анализа резистентного крахмала. В 2000 г. стандартизированный тест для определения содержания RS в продуктах питания был одобрен AACC International (9). Цифры, приведенные в этой статье, были определены этим методом (10).
Если бы приготовленные бобовые содержали столько стойкого крахмала, сколько сырые бобовые, нам было бы очень трудно их переваривать. Как и все продукты с высоким содержанием крахмала, при приготовлении бобовых в кипящей воде большие части кристаллических областей разрушаются, что снижает количество устойчивого крахмала. Но поскольку в бобовых так много амилозы, при варке разрушается меньшее количество устойчивого крахмала, чем в других продуктах, потому что некоторые формы кристаллической амилозы стабильны даже в кипящей воде (8).
- Полностью приготовленные бобовые содержат только около 4-5% от их общего веса (в пересчете на сухое вещество) в виде устойчивого крахмала, независимо от того, как долго они варятся.
- Охлаждение приготовленных бобовых до 24 часов в холодильнике увеличивает уровень устойчивого крахмала до 5-6% от общего веса (в пересчете на сухой вес), позволяя некоторым молекулам крахмала перекристаллизоваться. Целые консервированные бобы содержат примерно такое же количество, как и консервированные жареные бобы (10).
Может показаться, что это не очень стойкий крахмал, но он все же в 4-5 раз выше, чем в других крахмалистых продуктах, таких как белый хлеб и картофель (см. Таблицу ниже).Этого, безусловно, достаточно, чтобы оказать значительное влияние на образование SCFA, гликемический индекс, снижение инсулинового ответа, насыщения и калорийности (11). В общем, бобовые — это здоровый выбор не только из-за высокого содержания белка и других питательных веществ, но и потому, что они содержат одни из самых высоких уровней устойчивого крахмала среди всех продуктов питания.
| Продукты питания | Крахмал * | RS * | Гликемический индекс ** (12) |
| Картофель, запеченный | 94% | <1% | 93 |
| Белый хлеб | 88% | 1% | 70 |
| Фасоль черная, вареная | 60% | 5% | 30 |
| Фасоль пинто, вареная | 61% | 5% | 39 |
- Массовый процент в пересчете на сухое вещество
** Гликемический индекс ниже 55 считается низким показателем G.I. еда (12).
1. Национальная база данных по питательным веществам Министерства сельского хозяйства США для справочной информации (2011 г.).
2. Энглист, Х. Н., Кингман, С. М., и Каммингс, Дж. Х., Классификация и измерение важных для питания фракций крахмала. Евро. J. Clin. Nutr. 1992; 46: 533-550.
3. Кросби Г. А. Устойчивый крахмал делает углеводы лучше. Функциональные продукты питания и нутрицевтики, 2003; 6: 34-36.
4. Топпинг Д. Л. и Клифтон П. М. Короткоцепочечные жирные кислоты и функция толстой кишки человека: роль резистентных крахмальных и некрахмальных полисахаридов. Physiol. Rev.2001; 81 (3): 1031-1064.
5. Бехолл К. М. и Хоу Дж. К. Вклад клетчатки и устойчивого крахмала в метаболизируемую энергию. Являюсь. J. Clin. Nutr. 1995; 62: 1158S-1160S.
6. Raban, A., et al. Резистентный крахмал: влияние на постпрандиальную гликемию, гормональный ответ и чувство насыщения.Являюсь. J. Clin. Nutr. 1994; 60: 544-551.
7. Bednar, G.E., et al. Фракции крахмала и клетчатки в выбранных пищевых и кормовых ингредиентах влияют на их переваримость и ферментируемость в тонком кишечнике, а также на ферментируемость толстого кишечника in vitro на модели собак. J. Nutr. 2001; 131: 276-286.
8. Томпсон Д. Б. Стратегии производства устойчивого крахмала. Trends Food Sci. Tech. 2000; 11 (7): 245-253.
9. Переа А., Меда В.и Тайлер, Р. Т. Устойчивый крахмал: обзор аналитических протоколов для определения содержания устойчивого крахмала в пищевых продуктах. Food Res. Междунар. 2010; 43: 1959–1974.
10. Фаббри А. Д. Т., Шахт Р. В. и Кросби Г. А. Оценка содержания устойчивого крахмала в вареной черной фасоли, фасоли пинто и нуте. NFS Journal 3. 2016; 8-12.
11. Noah, L. et al. Переваривание углеводов из белой фасоли (Phaseolus vulgaris L.) у здоровых людей. J. Nutr.1998; 128: 977-985.
12. Бранд-Миллер, Дж., Волевер, Т. М. С., Колагиури, С., и Фостер-Пауэлл, К. Глюкозная революция: авторитетное руководство по гликемическому индексу. Marlowe & Co., NY, 1999.
СПРАВОЧНАЯ ИНФОРМАЦИЯ: Зерна крахмала находятся в хлоропласты высших растений и если листья, содержащие крахмал, держать в темноте в течение умеренного времени зерна крахмала будут пропадать.Если эти лишенные солнца листья подвергаются воздействию света, крахмал снова появляется в хлоропластах. Крахмал — первый видимый продукт фотосинтез. |