Гамма 7 01 комбайн: кухонный комбайн ЛЕПСЕ «Гамма-7-01» комплект 1 Киров Лепсе Сделано в России
Руководство по использованию кухонных комбайнов Лепсе — Олимп
Кухонный комбайн Лепсе гамма 7-01 – основные возможности
Это многофункциональный кухонный комбайн отечественного производства, который не уступает агрегатам от более известных брендов по своей функциональности и надежности. Используя кухонную машину, Гамма 7-01 Лепсе, можно легко и быстро:
- Приготовить фарш из мяса, рыбы или грибов.
- Натереть или очень тонко нарезать пластинками сыр, овощи, фрукты, шоколад, орешки.
- Приготовить мясо для беф-строганоф.
- Измельчить кофейные зерна (инструкция предусматривает регулировку степени измельчения кофейных зерен).
- Приготовить печенье замысловатой фигурной формы.
- Тонко раскатать тесто для приготовления пельменей.
Как показывают отзывы, кухонный комбайн гамма 7 Лепсе является незаменимым помощником на каждой кухне, особенно в период заготовок.
Кухонный комбайн Гамма 7-01 Лепсе – преимущества
Основное преимущество этого кухонного комбайна отечественного производства – это его многофункциональность.
Руководство кухонного комбайна гамма 7-01 Лепсе открывает перед пользователями информацию о том, что овощерезка в этом агрегате характеризуется особой конструкцией, благодаря которой в процессе работы диски измельчителя автоматически освобождаются т уже измельченных продуктов, благодаря чему кухонный комбайн может работать без остановки. Кроме того, инструкция этого агрегата содержит рекомендации, согласно которым хозяйка может в дополнение к основной кухонной машине приобрести дополнительно шнековую соковыжималку. Применение этой соковыжималки весьма разнообразно. Например, ее можно использовать не только для приготовления соков, но и для переработки ягод без косточки и томатов в пюре.
Руководство по эксплуатации этого кухонного комбайна содержит полую информацию о том, какие именно насадки входят в комплект, а также как правильно их использовать. Интересно, что с 2003 года кухонный комбайн Гамма 7-01 Лепсе комплектуется ножами для измельчения мяса, изготовленными из особой стали. Такой подход позволяет получить сочный фарш с минимальной потерей сока с волокон.
Лепсе предлагает хозяйкам не только качественный кухонный комбайн. В ассортименте продукции от этого производителя есть и электрическая мощная мясорубка Гамма 7-01-02.
Сервисное обслуживание кухонного комбайна Гамма от Лепсе
Несмотря на то, что кухонный комбайн, как и мясорубка 7-01-02 разработаны таким образом, чтобы их применение было максимально удобным и продуктивным, со временем некоторые запчасти изнашиваются. Как показывают отзывы хозяек, наиболее часто во время частого использования агрегата изнашиваются шестеренки. Кроме того, со временем может потребоваться замена ножа (если не удастся его подострить), или гайки-держателя решетки.
Запчасти на кухонный комбайн Лепсе гамма 7-01 найти сегодня не проблема. Запчасти и комплектующие к кухонному оборудованию этого типа продаются практически в каждом специализированном интернет магазине. Однако чтобы кухонный комбайн с новой комплектующей деталью работал как прежде, руководство к этому агрегату рекомендует покупать только «родные запчасти» от производителя, а не искать похожие по форме и размеру запчасти от других фирм.
Инструкция Мясорубки ЛЕПСЕ Гамма-7-01 (комплект 1)
Найденные инструкции для ЛЕПСЕ Гамма-7-01 (комплект 1)
В случае если инструкция не полная или нужна дополнительная информация по этому устройству,
если вам нужны дополнительные файлы: драйвера,
дополнительное руководство пользователя (производители зачастую для каждого продукта делают несколько различных документов технической помощи и руководств),
свежая версия прошивки, то вы можете задать вопрос
администраторам или всем пользователям сайта, все постараются оперативно отреагировать на ваш запрос и как можно быыстре помочь.
Ваше устройство имеет характеристики: Мощность: номинальная — 130 Вт, Производительность: 0.6 кг/мин, Максимальное время непрерывной работы: 30 мин, Перфорированный диск для фарша: есть, Насадка-терка: есть, Насадка для шинковки: есть, полные характеристики смотрите в следующей вкладке.
Скачать инструкцию к Мясорубки ЛЕПСЕ Гамма-7-01 (комплект 1) | |||
| ЛЕПСЕ_Гамма-7-01_(комплект_1)_0.pdf | Руководство пользователя | ||
| ЛЕПСЕ_Гамма-7-01_(комплект_1)_1.pdf | Скачать сертификат соответствия | ||
| Скачать Сообщить о нерабочей ссылке | |||
Полезные файлы и ПО
Для многих товаров, для работы с ЛЕПСЕ Гамма-7-01 (комплект 1) могут понадобиться различные дополнительные файлы: драйвера, патчи, обновления, программы установки.
Вы можете скачать онлайн эти файлы для конкретнй модели ЛЕПСЕ Гамма-7-01 (комплект 1) или добавить свои для бесплатного скачивания другим посетителями.
| Файлов не найдено |
| Добавить файл |
Инструкции для похожих Мясорубок
Если вы не нашли файлов и документов для этой модели то можете посмотреть интсрукции для похожих товаров и моделей, так как они зачастую отличаются небольшим изменениями и взаимодополняемы.
Отзывы о ЛЕПСЕ Гамма-7-01 (комплект 1)
Обязательно напишите несколько слов о преобретенном вами товаре, чтобы каждый мог ознакомиться с вашим отзывом или вопросом. Проявляйте активность что как можно бльше людей смогли узнать мнение настоящих людей которые уже пользовались ЛЕПСЕ Гамма-7-01 (комплект 1).
Характеристики ЛЕПСЕ Гамма-7-01 (комплект 1)
Текст описываающий харакетристики устройства.
| Технические характеристики | |
| Мощность | номинальная — 130 Вт |
| Производительность | 0.6 кг/мин |
| Максимальное время непрерывной работы | 30 мин |
| Насадки | |
| Перфорированный диск для фарша | есть |
| Насадка-терка | есть |
| Насадка для шинковки | есть |
| Прочие насадки | для нарезки печенья |
| Особенности | |
| Материал лотка | пластик |
| Материал корпуса | пластик |
| Дополнительная информация | жерновая приставка-кофемолка |
Поломки у Мясорубок
Здесь представлен список самых частых и распространенных поломок и неисправностей у Мясорубок. Если у вас такая поломка то вам повезло, это типовая неисправность для ЛЕПСЕ Гамма-7-01 (комплект 1) и вы можете задать вопрос о том как ее устранить и вам быстро ответят или же прочитайте в вопросах и ответах ниже.
Если у вас такая поломка то вам повезло, это типовая неисправность для ЛЕПСЕ Гамма-7-01 (комплект 1) и вы можете задать вопрос о том как ее устранить и вам быстро ответят или же прочитайте в вопросах и ответах ниже.
| Название поломки | Описание поломки | Действие |
|---|---|---|
| Не включается | вопрос | |
| Плохо рубит | вопрос | |
| Пахнет гарью, палеными проводами | вопрос | |
| Сильно шумит | вопрос | |
| Надрыв ремня | вопрос | |
| Поломка пластиковой шестерни | вопрос | |
| сильно шумит | вопрос | |
| Реверс | Реверс не работает | вопрос |
| не режит | сильно шумит и не режит | вопрос |
| некрутит крутит в пустую | вопрос | |
| неразбирается | вопрос | |
| НЕ крутит при нагрузке | При нагрузке не крутит | вопрос |
| не крутится шнек | двигатель работает. а шнек не крутится а шнек не крутится | вопрос |
| сломался шнек | вопрос | |
| Неправильно вставлен шнек и теперь не высовываться | вопрос | |
| Как фиксируется шнек при работе с насадкой для томатного сока | Шнек мясорубки и насадки двигаются по горизонтали на4-5 мм | вопрос |
| пластиковая втулка | пластиковая втулка | вопрос |
| шнек не вращается обратно | при нажатии на кнопку «реверс» ничего не происходит? | вопрос |
Поломки у ЛЕПСЕ Гамма-7-01 (комплект 1)
Если ни одна поломка из списка выше не подходит под описание для вашего случая то вы можете добавить свою поломку и мастера из сервисных центров или просто посетители сайта смогут вас проконсультруют. Это Бесплатно!
Добавить поломку
Сервисы специалзирующиеся на ремонте Мясорубок
В нашей базе сейчас зарегестрированно 18 353 сервиса в 513 города России, Беларусии, Казахстана и Украины.
Сервисы выбранные пользователями
Сервисы по порядку
загрузть ещеинструкция по применению на видео, отзывы
Бытовая техника создается для того, чтобы сделать жизнь человека комфортной. Процесс приготовления пищи – не исключение. Его можно облегчить и приятно разнообразить, если приобрести хорошее оборудование для обработки продуктов. Давайте далее в этой статье мы расскажем все про кухонные комбайны Гамма: виды, эксплуатация, достоинства.
Выбираем кухонный комбайн Гамма
Краткое описание
Кухонный комбайн Гамма 7-01 – это чудесный помощник для хозяек, которые любят готовить интересные блюда своими руками. Такой агрегат обеспечивает пользователю широкие возможности касательно обработки продуктов. Он характеризуется низким энергопотреблением и имеет несколько режимов работы.
Такая электромясорубка удобна в использовании, а мыть ее после этого совсем несложно. И отзывы довольных покупательниц о применении Лепсе — тому прекрасное подтверждение.
Достоинства
Отметим, что кухонный комбайн Лепсе Гамма 7-01 обладает высокими эксплуатационными параметрами. Многие пользователи утверждают, что эта марка создает действительно удачные по соотношению цены-качества модели кухонных комбайнов и мясорубок. И если разбираться в этом вопросе, то пройти мимо такого оборудования просто невозможно. Ведь это и мясорубка, и комбайн, и кофемолка в одном изделии.
Основными преимуществами, которые характерны для кухонных комбайнов Гамма Лепсе 7, называют следующие качества:
- высокая функциональность обеспечивается отменным качеством сборки и долговечными запчастями;
- практичность: применение такого оборудования в домашних условиях не отличается какой-либо сложностью;
- долгий срок службы: Гамма 7-01, 3 и 7, крайне редко ломаются, а если это и произошло, всегда можно заменить пришедшие в негодность запчасти;
- красивый внешний вид изделий Лепсе;
- доступная стоимость;
- понятная инструкция по эксплуатации.


Запчасти
С 2003 года модели электромясорубки этого бренда комплектуется инновационным ножом, который изготавливается из высокопрочной стали. Качество такого ножа обеспечивает возможность без лишних усилий готовить мясной фарш без потери натурального сока из волокон.
Также отметим, что Лепсе 7 отличаются от многих схожих по стоимости агрегатов тем, что в этих кухонных комбайнах присутствует жерновая приставка-кофемолка. Она позволяет перемолоть кофе так однородного, что напиток получается, по истине, королевским.
А при желании пользователь в дополнение к кухонному комбайну Лепсе 7-01, 3 и 7 можно купить и шнековую соковыжималку «Гамма-СВ», которая предназначается для переработки ягод и томатов в сок-пюре. Это отличное дополнение к основной модели, способная обеспечить вас вкусными и, главное, полезными десертами.
Особенности использования
Инструкция для кухонного комбайна Гамма очень проста для понимания, поэтому научиться пользоваться таким оборудованием для кухни очень просто.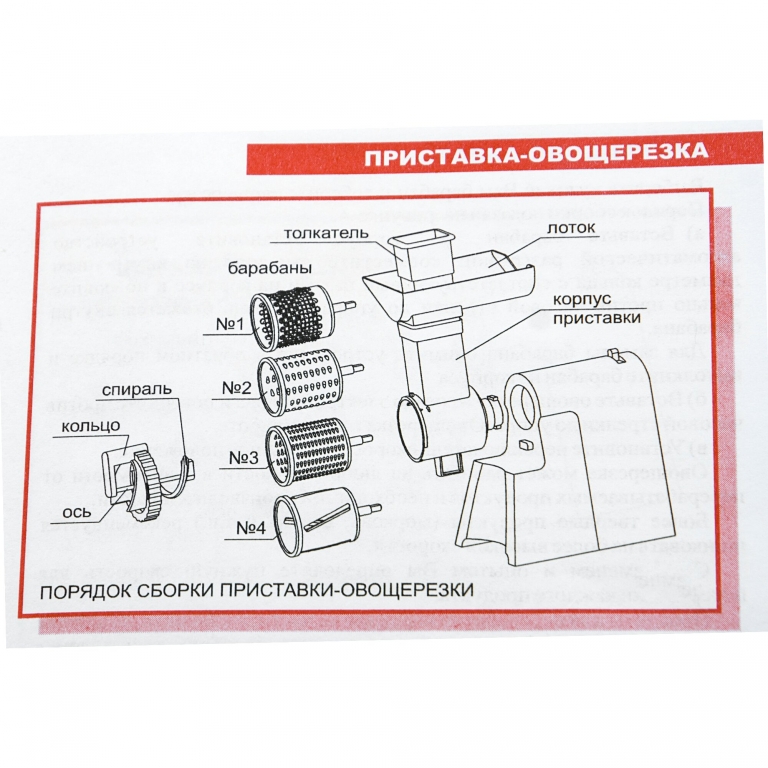 Ее может освоить даже подросток. Тем более что польза от применения кухонного комбайна этого бренда весьма очевидна.
Ее может освоить даже подросток. Тем более что польза от применения кухонного комбайна этого бренда весьма очевидна.
Чтобы понять, действительно ли говорят правду отзывы о продукции Гамма в интернет сети, вам нужно будет всего лишь раз попробовать его в эксплуатации.
Вам поможет высококачественный и доступный по стоимости кухонный комбайн Гамма 7. Воспользуйтесь им, если вы прочили инструкцию пользователя и решили выполнить следующие операции своими руками:
- сделать фарш из говядины, свинины, баранины или рыбы без лишних проблем и хлопот. Причем нарезка мясопродуктов может быть как мелкой, так и крупной. То есть, самый обычный кухонный комбайн данной марки выступает в роли эффективной мясорубки, что весьма удобно в быту;
- подготовить овощи для салата или рагу. Он натрет, нашинкует или тонко нарежет продукты за считанные минуты, а пользователь при этом не потратит своих сил. Также такое оборудование легко справляется с нарезкой фруктов, сыра и других продуктов;
- измельчить шоколад или орехи до мукообразного состояния или измолоть зерна кофе, чтобы получить вкуснейший напиток;
- порадовать близким блюдом типа «беф-строганов» с нежной консистенцией;
- приготовить тесто разных видов для создания выпечки или пельменей.

Высокая функциональность продукции Гамма выделяет эти модели на фоне аналогичной по стоимости продукции иных брендов. К тому же повторимся: запчасти для кухонного комбайна Гамма всегда присутствуют в продаже и характеризуются низкой стоимостью, что очень удобно и продлевает срок службы такой техники.
А видео в конце этой статьи продемонстрирует кухонный комбайн Гамма в действии. Ознакомьтесь с ним, а также инструкцией к модели, чтобы ее использование не омрачалось трудностями.
Загрузка…№ | наименование | внешний вид | характеристики |
| 1 | вид установки шестерней электромясорубок ГАММА 7-01 (г.Киров) | вид установки вид сзади вид сбоку вид сверху | |
| 2 | втулка уплотнительная для электромясорубки ГАММА (г.  Киров) Киров) | вид спереди вид cзади | •ширина-12 мм; •Ø внешний-35 мм. |
| 3 | манжета уплотнительная для мясорубки ГАММА (г.Киров) | вид спереди вид cзади | •ширина-6,5 мм; •Ø 21 мм. |
| 4 | шестерня для мясорубки ЛЕПСЕ (г.Киров) | вид спереди вид cзади | •длина-42 мм; •вес-44 г; •Ø 30 мм/51 мм. |
| 5 | шестерня для электромясорубки ГАММА (г.Киров) | вид спереди вид cбоку вид cзади | •Ø широкой части-82 мм; •количество зубьев (косозубые) в широкой части-96 штук; •толщина широкой части -11 мм; •Ø узкой части-20 мм; •количество зубьев (косозубые) в узкой части-9 штук; •толщина шестерни -33 мм. |
| 6 | держатель втулки для мясорубки ГАММА (г.Киров) | вид спереди вид cбоку вид cзади | •длина-53 мм; •вес-64 г; •Ø 21 мм/62 мм.  |
| 7 | шнек для электромясорубки ЛЕПСЕ (г.Киров) | шнек для мясорубки | •длина-126 мм. |
| 8 | втулка для мясорубки ЛЕПСЕ (г.Киров) | вид спереди вид cбоку вид cзади | •длина-35 мм; •вес-14 г; •Ø 22 мм. |
| 9 | нож для мясорубки ГАММА (г.Киров) | вид спереди вид cзади | •подходит для большинства отечественных электромясорубок и ручных мясорубок; •толщина-8 мм; •Ø 48 мм. |
| 10 | решётка для мясорубки ГАММА (г.Киров) (для приготовления бефстроганов) | решётка мясорубки для бефстроганов | •подходит для большинства отечественных электромясорубок и ручных мясорубок; •толщина-5 мм; •Ø 54 мм. |
| 11 | двигатель к мясорубке ГАММА 7-01 (г.  Киров) Киров) | двигатель к мясорубке | •для мясорубок ГАММА 7-01. |
| 12 | корпус к мясорубке ГАММА 7-01 (г.Киров) | корпус к мясорубке вид внутри | •для мясорубок ГАММА 7-01. |
| 13 | лоток шинковки мясорубки ЛЕПСЕ 7-01 (г.Киров) | лоток шинковки вид снизу | •для мясорубок ЛЕПСЕ,ГАММА; •габаритные размеры: 141×177 мм; •посадочный размер: 54×59 мм. |
| 14 | шнур питания к мясорубке | шнур питания | •для мясорубки. |
| 15 | переключатель к мясорубке ГАММА 7-01 (г.Киров) | корпус к мясорубке | •для мясорубок ГАММА 7-01. |
| 16 | крышка корпуса нижняя для мясорубки ГАММА 7-01 (г.Киров) | крышка корпуса нижняя | •для мясорубок ГАММА 7-01.  |
| 17 | насадка для мясорубки ГАММА (насадка овощерезка) (г.Киров) | насадка овощерезка | •для мясорубки: ГАММА; •три барабана: 1.для мелкого шинкования, 2.крупного шинкования, 3.перетирания. |
| 18 | насадка для мясорубки ГАММА (насадка для сока) (г.Киров) | насадка для сока | •для мясорубок: ГАММА. |
Отзыв про Мясорубка Лепсе «Гамма-7-01»: «Все детали хлипкие, слабая мясорубка»
Давно хотели электрическую мясорубку, но прикинули, что комбайн повыгоднее будет по функциям, заготовок делаю массу, да и просто готовить люблю. Приценивались, смотрели и обратили внимание на это, как нам тогда показалось, чудо- техники. Привлекло, что сделано в России, в славном Кирове и цена. К тому же, покупали ее в небольшом отделе, продавец уж очень ее хвалила. Понимаю, это ее хлеб, но она была убедительна, плюс, говорила, что на этот комбайн не бывает возвратов, такие они качественные.
Цена зависела от сборки, какую комплектацию выберешь. Количество насадок можно было менять, например, не нужна кофемолка- не бери.
Комплектность полная: мясорубка, овощерезка, кофемолка, соковыжималка, насадка для профилирования теста. Мы не брали кофемолку и насадку для теста.
А теперь о использовании и качестве, собственно говоря.
Начали мы с овощерезки: терка, соломка, шинковка. Тут претензий к комбайну не было долгое время. Пока не попалась нам очень твердая морковь. Может старовата была, может что то неудачно сложилось, но одна из насадок просто погнулась. Резать на ней еще можно, но все же, уже не то.
Погнувшаяся насадка
Далее, соковыжималка. Там такая сменная решетка
сменная решетка
, которая очень легко гнется и потом ее очень трудно вернуть в первоначальный вид, достал, помыл, а вставить- начинаешь изгаляться. Видимо, к этому делу нужно приспособиться, потому что как я не следовала инструкции, у меня все равно забивало решетку, заклинивало нож- шнек, приходилось его разбирать каждые две минуты и чистить. Не соковыжимание, а мучение. В защиту хочу сказать, что видимо, тут нужно как то приноравливаться, видимо, просто у меня не получилось с этой насадкой подружиться. Но сколько нужно намучатся, что бы это сделать я не знаю.
Не соковыжимание, а мучение. В защиту хочу сказать, что видимо, тут нужно как то приноравливаться, видимо, просто у меня не получилось с этой насадкой подружиться. Но сколько нужно намучатся, что бы это сделать я не знаю.Наконец, самое главное, мясорубка. Фарш получался неоднородный, приходилось перекручивать несколько раз. Что то забивалось, приходилось вечно раскручивать.
И очень скоро, нам пришлось отдать шнек на ремонт. При перекручивании мяса он застрял, неловко провернулся и у той части, которая вставляется в сам комбайн, согнуло грани (на фотографии обвела желтым). Вытаскивали- вытаскивали и отдали в ремонт.
Нам сменили комбайн полностью и мы решили измельчить печенье, теперь шнек погнуло с той стороны, где вставлялся нож (отмечено красным), он опять внезапно застрял. Хотя все открывалось и чистилось в процессе работы.
шнек
Плюнули на нее и поставили в шкаф. Достала только ради отзыва. Планирую пользоваться овощерезкой и только.
комбайн
Гамма-01
Лицензионное соглашение на использование программного обеспечения «Гамма-Поток»
Перед использованием программы, пожалуйста, ознакомьтесь с условиями нижеследующего лицензионного соглашения.
Любое использование Вами программы означает полное и безоговорочное принятие Вами условий настоящего лицензионного соглашения.
Если Вы не принимаете условия лицензионного соглашения в полном объёме, Вы не имеете права использовать программу в каких-либо целях.
1. Общие положения
1.1. Настоящее Лицензионное соглашение («Лицензия») устанавливает условия использования программного обеспечения «Гамма-Поток» («Программу») и заключено между любым лицом, использующим Программу («Пользователь») и ООО «НПО ПАС», Россия, 109129, г. Москва, 8-я ул. текстильщиков, д.18, корп. 3, являющимся правообладателем исключительного права на Программу («Правообладатель»).
1.2. Копируя Программу, устанавливая её на свое устройство или используя Программу любым образом, Пользователь выражает свое полное и безоговорочное согласие со всеми условиями Лицензии.
1.3. Использование Программы разрешается только на условиях настоящей Лицензии. Если Пользователь не принимает условия Лицензии в полном объёме, Пользователь не имеет права использовать Программу в каких-либо целях. Использование Программы с нарушением (невыполнением) какого-либо из условий Лицензии запрещено.
Использование Программы с нарушением (невыполнением) какого-либо из условий Лицензии запрещено.
1.4. Использование Программы Пользователем на условиях настоящей Лицензии осуществляется безвозмездно. Использование Программы на условиях и способами, не предусмотренными настоящей Лицензией, возможно только на основании отдельного соглашения с Правообладателем.
1.5. К настоящей Лицензии и всем отношениям, связанным с использованием Программы, подлежит применению право Российской Федерации и любые претензии или иски, вытекающие из настоящей Лицензии или использования Программы, должны быть поданы и рассмотрены в суде по месту нахождения Правообладателя.
1.6. Правообладатель может предоставить Пользователю перевод настоящей Лицензии с русского на другие языки, однако в случае противоречия между условиями Лицензии на русском языке и ее переводом, юридическую силу имеет исключительно русскоязычная версия Лицензии.
2. Права на программу
2.1. Исключительное право на Программу принадлежит Правообладателю.
3. Права Пользователя
3.1. Правообладатель безвозмездно, на условиях простой (неисключительной) лицензии, предоставляет Пользователю непередаваемое право использования Программы на территории всех стран мира указанным в настоящем разделе способами.
3.2. Пользователь вправе применять Программу по прямому функциональному назначению в соответствии с Руководством Пользователя, в целях чего произвести установку Программы на настольном персональном компьютере или ином устройстве (далее «Устройствах»).
В соответствии с настоящим пунктом Пользователь вправе произвести установку Программы на неограниченное число Устройств, а также хранить или установить копии Программных продуктов на устройстве хранения данных (например, на сервере сети), предназначенном и используемом исключительно для установки Программы на других имеющихся компьютерах в пределах локальной сети.
3.3. Пользователь вправе воспроизводить Программное обеспечение «Гамма-Поток», экземпляры которого распространяются Правообладателем, в некоммерческих целях на безвозмездной основе на любых Устройствах и прочих видах материальных носителей при условии сохранения в неизменном виде состава и содержания Программы по сравнению с тем, как они предоставляются и/или рекомендуются для использования Правообладателем.
3.4. Пользователь вправе распространять Программу посредством передачи ее экземпляров, воспроизведенных в соответствии с пунктом 4.3 настоящего Соглашения, в некоммерческих целях на безвозмездной основе и при соблюдении их целостности, если иное не предусмотрено письменным разрешением Правообладателя.
4. Ограничения
4.1. За исключением использования в объёмах и способами, прямо предусмотренными настоящей Лицензией или законодательством РФ, Пользователь не имеет права изменять, декомпилировать, дизассемблировать, дешифровать и производить иные действия с объектным кодом Программы, имеющие целью получение информации о реализации алгоритмов, используемых в Программе, создавать производные произведения с использованием Программы, а также осуществлять (разрешать осуществлять) иное использование Программы, любых компонентов Программы, хранимых Программой на устройстве Пользователя, без письменного согласия Правообладателя.
4.2. Пользователь не имеет право воспроизводить и распространять Программу в коммерческих целях (в том числе за плату), в том числе в составе сборников программных продуктов, без письменного согласия Правообладателя.
4.3. Пользователь не имеет права распространять Программу в виде, отличном от того, в котором он её получил, без письменного согласия Правообладателя.
4.4. Программа должна использоваться (в том числе распространяться) под наименованием: «Гамма-Поток». Пользователь не вправе изменять и/или удалять наименование Программы, знак охраны авторского права (copyright notice) или иные указания на Правообладателя.
5. Условия использования отдельных функций программного обеспечения «Гамма-Поток»
5.1. Выполнение некоторых функций Программы (регистрация, обновление) возможно при наличии доступа к сети Интернет. Пользователь самостоятельно получает и оплачивает такой доступ на условиях и по тарифам своего оператора связи или провайдера доступа к сети Интернет.
5.2. Технические данные применяемого в программе оборудования производства ООО «НПО ПАС», а также иные объекты интеллектуальных прав, доступ к которым предоставляет Программа (далее — Данные), принадлежат Правообладателю и/или их владельцам, указанным в пункте 10 настоящей Лицензии, и предназначены исключительно для использования Пользователем в рамках функциональных возможностей Программы.
5.3. Программа предоставляет Пользователю функциональную возможность расчета массы газовых огнетушащих веществ, построения трубных разводок и проведения гидравлических расчетов, указывающих время выпуска газовых огнетушащих веществ и распределение их в защищаемых зонах (направлениях). Ввод данных для расчетов, графическое построение трубных разводок (систем пожаротушения) не модифицирует исходные Данные и допускается исключительно для удобства использования Пользователем Программы.
Пользователь самостоятельно и всецело несёт все риски и ответственность за правильность введенных данных, построение трубных разводок. Правообладатель не гарантирует правильность, оптимальность, актуальность и/или соответствие трубных разводок действующим нормам и правилам и не несет ответственности за выбор Пользователем огнетушащих веществ, и конфигурации трубных разводок.
6. Ответственность по Лицензии
6.1. Программа (включая Данные) предоставляется на условиях «как есть» (as is). Правообладатель не предоставляет никаких гарантий в отношении безошибочной и бесперебойной работы Программы или отдельных её компонентов и/или функций, соответствия Программы конкретным целям и ожиданиям Пользователя, не гарантирует достоверность, точность, полноту и своевременность Данных, а также не предоставляет никаких иных гарантий, прямо не указанных в настоящей Лицензии.
6.2. Правообладатель не несет ответственности за какие-либо прямые или косвенные последствия какого-либо использования или невозможности использования Программы (включая Данные) и/или ущерб, причиненный Пользователю и/или третьим сторонам в результате какого-либо использования, неиспользования или невозможности использования Программы (включая Данные) или отдельных её компонентов и/или функций, в том числе из-за возможных ошибок или сбоев в работе Программы, за исключением случаев, прямо предусмотренных законодательством.
6.3. Ни при каких условиях, включая, но, не ограничиваясь, невнимательностью или небрежностью пользователей, компания ООО «НПО ПАС» не несет ответственности за любой ущерб (прямой или косвенный, случайный или закономерный), включая, но не ограничиваясь, потерей данных или прибылей, связанной с использованием или невозможностью использования этой программы. В случае, если использование данной программы приведет к необходимости дополнительного обслуживания, исправления или ремонта вашего оборудования или восстановления данных, все связанные с этим затраты оплачиваются Вами.
7. Обновления/новые версии программного обеспечения
7.1. Действие настоящей Лицензии распространяется на все последующие обновления/новые версии Программы. Соглашаясь с установкой обновления/новой версии Программы, Пользователь принимает условия настоящей Лицензии для соответствующих обновлений/новых версий Программы, если обновление/установка новой версии Программы не сопровождается иным лицензионным соглашением.
8. Политика конфиденциальности
8.1. Порядок обработки и защиты информации о Пользователях, полученной Право-обладателем в связи с регистрацией Пользователями Программы определяется Политикой конфиденциальности с учетом особенностей, установленных настоящим Соглашением.
8.2. Правообладатель собирает и обрабатывает персональную информацию в следующих целях:
8.2.1. Формирование и предоставление Пользователю регистрационного ключа;
8.2.2. Предоставление пользователю технической поддержки;
8.2.3. Уведомление при появлении новых версий программы.
8.3. В отношении персональной информации Пользователя сохраняется ее конфиден-циальность кроме случаев предусмотренных российским или иным применимым законодательством в рамках установленной законодательством процедуры.
9. Изменение условий настоящей Лицензии
9.1. Настоящее лицензионное соглашение может изменяться Правообладателем в одностороннем порядке. Уведомление Пользователя о внесенных изменениях в условия настоящей Лицензии публикуется на официальном сайте в сети интернет. Указанные изменения в условиях лицензионного соглашения вступают в силу с даты их публикации, если иное не оговорено в соответствующей публикации.
10. Информация о правообладателях и обладателях лицензии
10.1. Методика гидравлического расчета установок газового пожаротушения (ПАС 725.00.000.РР) – © ООО «НПО Пожарная автоматика сервис» (http://www.npo-pas.com)
10.2. Программное обеспечение «Гамма-Поток» – © ООО «НПО Пожарная автоматика сервис» (http://www.npo-pas.com)
Отзывы пользователей
«Спасибо за регистрацию. Хочу оставить свой положительный отзыв о ПО «Гамма-Поток». Очень доволен возможностью самостоятельно выполнять расчеты. С данным программным продуктом можно не только получить расчет по конкретному объекту, но и в целом лучше вникнуть в саму методику гидравлического расчета и расчета массы ГОТВ. Программа очень гибкая по своему функционалу, на любой стадии расчета можно менять исходные данные(есть правда маленький недочет: на WIN7 при вводе названия помещения графа с температурой Тм не отображается. Она есть, но почему-то не отображается). Построение аксонометрической схемы (! и ее последующее редактирование) — это просто песня, я доволен. Вообщем, мой выбор, как проектировщика, в пользу оборудования НПО ПАС однозначный. Моя благодарность руководству компании и разработчикам ПО «Гамма-Поток»!»
Ведущий проектировщик «Институт промышленной и пожарной безопасности» г. Новокузнецк
«Благодарим за отличный программный продукт.»
Проектная группа ООО «ИнжПожПроект»
«Большое спасибо! Наконец-то стало возможным экономить свое время!»
Проектировщик ООО «ПожСтройПроект»
«Программа удобна в использовании. Исходные данные вводить удобно. Все удобно, появляются подсказки, если что-то выполнено неправильно.»
Проектировщик ООО «Проектное Бюро»
«Огромное спасибо за продукт! Всё вполне функционально и доступно для понимания с первой минуты пользования программой. Из пожеланий: хотелось бы иметь возможность привязки трубной разводки к защищаемому помещению, то есть рисовать разводку на подложке dwg файлов.»
Проектировщик ОАО «ЧЕЛЯБСПЕЦАВТОМАТИКА»
«Сложностей с работой в программе не возникло. Вот пару предложений:
— хотелось бы увидеть расчёт площади проема для сброса избыточного давления;
— хотелось бы аксонометрическую схему, выполненную в программе Гамма-Поток, сохранить в формате dwg.
А так программа ОТЛИЧНАЯ! СПАСИБО!»
Проектировщик ЗАО «ФИТТИХ-КЛЕН»
«Спасибо за качественный, удобный в работе продукт. Программа позволяет значительно экономить время и выгодно отличается от аналогичных программ, существующих на рынке. Большим преимуществом является, то что ПО «Гамма-Поток» предоставляется бесплатно.»
Проектная группа ООО «Фламакс»
Хаотическая динамика, применяемая к предсказанию энергии пьезоэлектрического комбайна с временной задержкой
Алдаир А., Алсаеди Э. (2020) Энергия регенерации для нелинейной активной системы подвески с использованием электромагнитного привода. Иракский J Electr Electron Eng 16 (2): 113–125
Статья Google Scholar
Behnia S, Rahimi F, Khodavirdizadeh M (2019) Управление светом в решетке фотонного резонатора: подход квантового хаоса.Opt Commun 446: 171–177
Статья Google Scholar
Бутусов Д.Н., Пестерев Д.О., Тутуева А.В., Каплун Д.И., Непомуцено Е.Г. (2021) Новый метод количественной оценки хаотической динамики, основанный на различиях между полунеявными схемами интегрирования. Commun Nonlinear Sci Numer Simul 92: 105467
MathSciNet Статья Google Scholar
Чакмак Болат Ф., Басаран С., Кара М. (2021) Исследование сбора энергии в композитных балках с разными углами ламинирования при динамических воздействиях.Compos Struct 270: 114056
Артикул Google Scholar
Chakrabarti A, Arora M (eds) (2019) Индустрия 4.0 и передовое производство: материалы I-4AM. Конспект лекций по машиностроению. Springer, Singapore, p 2021
Feng W, Li Q, Lu Q, Li C, Wang B (2021) Машина вектора групповой релевантности для локализации и реконструкции разреженных сил. Сигнальный процесс Mech Syst 161: 107900
Артикул Google Scholar
Fu Y, Ouyang H, Davis RB (2020) Влияние электрических свойств на вибрации через электромеханическую связь при сборе трибоэлектрической энергии. J Phys D Appl Phys 53 (21): 215501
Статья Google Scholar
Gao M, Wang Y, Wang Y, Yao Y, Wang P, Sun Y, Xiao J (2020) Моделирование и экспериментальная проверка четверной стабильной системы сбора энергии с дробным демпфированием для использования в беспроводных сенсорных сетях. Энергетика 190: 116301
Статья Google Scholar
Gopala Rao LVV, Narayanan S (2020) Оптимальный отклик модели полуавтомобиля с демпфером Sky-Hook на основе управления LQR. Int J Dyn Control 8 (2): 488–496
Статья Google Scholar
Хоссейннежад П., Бехниа С., Фатизаде С. (2020) Динамическая стабильность фотосинтетических систем: подход квантового хаоса. Солитоны хаоса Фракталы 139: 110279
MathSciNet Статья Google Scholar
Хуанг X, Ян Б. (2021) Улучшение сбора энергии из импульсных возбуждений с помощью нелинейно настраиваемого бистабильного сборщика энергии. Сигнальный процесс Mech Syst 158: 107797
Артикул Google Scholar
Kecik K (2021) Одновременное снижение вибрации и сбор энергии от поглотителя маятникового типа. Commun Nonlinear Sci Numer Simul 92: 105479
MathSciNet Статья Google Scholar
Кумар Р., Гупта С., Али С.Ф. (2019) Сбор энергии из хаоса в базовом возбужденном двойном маятнике. Сигнальный процесс Mech Syst 124: 49–64
Статья Google Scholar
Liu M, Mi J, Tai W-C, Zuo L (2021) Новая конфигурация для высокой выходной мощности и высокоэффективного сбора энергии вибрации. Appl Energy 295: 116957
Артикул Google Scholar
Liu M, Tai W-C, Zuo L (2021) Сбор энергии вибрации с использованием инерционной системы с двумя степенями свободы. Сигнальный процесс Mech Syst 146: 107000
Артикул Google Scholar
Маамер Б., Бугамура А., Фатх Эль-Баб А.М., Фрэнсис Л.А., Тунси Ф. (2019) Обзор усовершенствований конструкции и методов сбора механической энергии с использованием пьезоэлектрических и электромагнитных схем. Energy Convers Manage 199: 111973
Статья Google Scholar
Sah DK, Amgoth T (2020) Схемы сбора возобновляемой энергии в беспроводных сенсорных сетях: обзор. Inform Fusion 63: 223–247
Статья Google Scholar
Taghavifar H (2021) Новый подход к накоплению энергии для гибридной электромагнитной системы подвески внедорожников с учетом деформируемости местности. Обработка сигналов Mech Syst 146: 106988
Статья Google Scholar
Tan T, Wang Z, Zhang L, Liao W-H, Yan Z (2021) Пьезоэлектрический автопараметрический сбор энергии вибрации с функцией контроля хаоса. Сигнальный процесс Mech Syst 161: 107989
Артикул Google Scholar
Таварес Р., Рудерман М. (2020) Сбор энергии с использованием пьезоэлектрических преобразователей для подвесных систем. Мехатроника 65: 102294
Артикул Google Scholar
Touairi S, Bouzid A, Mabrouki M Дорожная обработка рекуперативных подвесок мотоциклов и сбор энергии, p 020017
Touairi S, Khouya Y, Bahanni C, Khaouch Z, Mabrouki M (2019) Мехатронное управление и моделирование Пьезоэлектрический привод. В: Международная конференция по беспроводным технологиям, встроенным и интеллектуальным системам (WITS), 2019 г. (Фес, Марокко, апрель 2019 г.), IEEE, стр. 1–6
Touairi S, Khouya Y, Bahanni C, Mabrouki M ( 2019) Скользящее управление пьезоэлектрическим приводом с использованием графа связи.В: 5-я Международная конференция по оптимизации и приложениям (ICOA), 2019 г. (Кенитра, Марокко, апрель 2019 г.), IEEE, стр. 1–7
Touairi S, Mabrouki M (2020) Мехатронное моделирование и управление рекуперацией энергии в мотоциклетных шинах. В: 2020 6-я международная конференция IEEE по оптимизации и приложениям (ICOA) (Бени Меллал, Марокко, апрель 2020 г.), IEEE, стр. 1–5
Touairi S, Mabrouki M (2020) Оптимизация харвестерной системы в встроенные автомобильные системы с помощью алгоритма моделирования Bond Graph.В: 2020 6-я международная конференция IEEE по оптимизации и приложениям (ICOA) (Бени Меллал, Марокко, апрель 2020 г.), IEEE, стр. 1–6
Touairi S, Mabrouki M (2021) Оценка управления и моделирования Пьезоэлектрическая система уборки урожая. Int J Dyn Control
Toyabur R, Salauddin M, Cho H, Park JY (2018) Мультимодальный гибридный комбайн для сбора энергии, основанный на пьезоэлектрических электромагнитных механизмах для низкочастотных колебаний окружающей среды. Energy Convers Manage 168: 454–466
Статья Google Scholar
Wang J, Su Z, Li H, Ding L, Zhu H, Gaidai O (2020) Создание эффекта следа для улучшения сбора чистой морской энергии за счет вибраций, вызванных потоком. Ocean Eng 208: 107455
Статья Google Scholar
Ван Р, Калнай Э, Балачандран Б (2019) Прогнозирование хаотической динамики на основе нейронных машин. Нелинейный Dyn 98 (4): 2903–2917
Статья Google Scholar
Ван X-S, Манн Б.П. Выбор аттрактора в нелинейном сборе энергии с использованием глубокого обучения с подкреплением 20
Вията А., ПолчиНСски К., Аврейцевич Дж. (2021) Теоретический и численный анализ регулярных односторонних колебаний в одиночной маятниковой системе, управляемой магнитным полем. Сигнальный процесс Mech Syst 150: 107229
Артикул Google Scholar
Xiong Q, Qin B, Li X, Zuo LA (2020) Управление демпфированием на основе правил подвески энергоемкого транспортного средства на основе MMR.В: 2020 American Control Conference (ACC) (Денвер, Колорадо, США, июль 2020 г.), IEEE, стр. 2262–2267
Zerimeche H, Houmor T, Berkane A (2020) Комбинированная синхронизация различных измерений дробно- упорядочить неавтономные хаотические системы с помощью матрицы масштабирования. Int J Dyn Control
Регулирование кормодобывания муравьев комбайном как замкнутая возбудимая система
Abstract
Колонии муравьев регулируют активность в ответ на изменение условий без использования централизованного контроля.Колонии муравьев-сборщиков пустынь собирают корм для семян и регулируют сбор корма, чтобы найти компромисс между расходами и получением воды. Фуражиры теряют воду, находясь на улице в сухом воздухе, но муравьи получают воду, метаболизируя жиры в семенах, которые они едят. Предыдущая работа показывает, что скорость, с которой уходящий собиратель покидает гнездо, зависит от его недавних кратких контактов антенн с приближающимися фуражирами, несущими корм. Мы исследуем, как этот процесс может обеспечить показатели кормодобывания, устойчивые к неопределенности и реагирующие на температуру и влажность в поминутной шкале времени.Чтобы изучить возможные механизмы, мы разработали аналитическую модель низкой размерности с небольшим количеством параметров, которая отражает наблюдаемое поведение при поиске пищи. Модель использует динамику возбудимости для представления реакции на взаимодействия внутри гнезда и случайное распределение задержки для представления времени поиска пищи за пределами гнезда. Мы показываем, как обратная связь от уходящих собирателей, возвращающихся в гнездо, стабилизирует входящие и исходящие нормы кормодобывания до общего значения, определяемого изменчивостью доступных фуражиров.Модель демонстрирует критическую изменчивость, выше которой кормодобывание продолжается с постоянной скоростью, а ниже которого кормление прекращается. Чтобы объяснить, как интенсивность кормодобывания приспосабливается к температуре и влажности, мы предлагаем собирателям изменять свою изменчивость после того, как они покидают гнездо и подвергаются воздействию окружающей среды. Наше исследование подчеркивает важность обратной связи в регулировании кормодобывающей деятельности и показывает, как модуляция изменчивости может объяснить, как кормодобывающая деятельность реагирует на условия и варьируется в зависимости от колонии.Наша модель объясняет роль обратной связи во многих временных масштабах в коллективном поведении и может быть обобщена на другие системы, управляемые возбудимой динамикой, такие как нейронные сети.
Информация об авторе
Мы исследуем коллективное поведение, которое позволяет колониям пустынных муравьев-собирателей регулировать добычу пищи в зависимости от условий окружающей среды. Мы разрабатываем аналитическую модель, объединяющую три процесса: 1) взаимодействия между фуражирами, возвращающимися в гнездо, и доступными фуражирами, ожидающими внутри гнезда, 2) влияние этих взаимодействий на вероятность того, что доступные фуражиры покинут гнездо на корм, и 3) возвращение фуражиров в гнездо после нахождения семян.Мы предлагаем механизм, с помощью которого доступные собиратели изменяют свою реакцию на взаимодействия после первого контакта с окружающей средой. Мы показываем, как это приводит к тому, что показатели кормодобывания в колониях приспосабливаются к условиям окружающей среды с течением времени от минут до часов. Наша модель может оказаться полезной для изучения других классов систем с динамикой возбудимости, которые демонстрируют как стабильность в поведении, так и гибкость по отношению к условиям окружающей среды.
Образец цитирования: Pagliara R, Gordon DM, Leonard NE (2018) Регулирование кормодобывания муравьев-комбайнов как возбудимой системы с замкнутым контуром.PLoS Comput Biol 14 (12): e1006200. https://doi.org/10.1371/journal.pcbi.1006200
Редактор: Джессика К. Флэк, Институт Санта-Фе, США
Поступила: 12.05.2018; Одобрена: 5 ноября 2018 г .; Опубликован: 4 декабря 2018 г.
Авторские права: © 2018 Pagliara et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Необработанные данные и сценарии моделирования модели MATLAB доступны в репозитории в DataSpace Принстонского университета по постоянному адресу: http://arks.princeton.edu/ark:/88435/dsp01qv33s040d.
Финансирование: Эта работа была частично поддержана грантом Управления военно-морских исследований США № N00014-14-1-0635 (NEL) и Фондом Уильяма и Флоры Хьюлетт (DMG). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Социальные колонии насекомых действуют без централизованного контроля. Колонии сохраняют согласованность и пластичность перед лицом возмущений и изменений, даже несмотря на то, что люди имеют ограниченную и неопределенную информацию о состоянии группы и состоянии окружающей среды. Коллективное поведение возникает из реакции людей на социальные взаимодействия и их оценки местной среды [1–4].Изучение социальных насекомых дает возможность исследовать открытые фундаментальные вопросы о том, как коллективное поведение приспосабливается к различным условиям и как небольшие различия в этих приспособлениях могут привести к большим различиям в поведении между группами.
Регулирование кормовой активности в колониях муравьев-сборщиков ( Pogonomyrmex barbatus ) является хорошо изученным примером коллективного поведения [5]. Муравьи-комбайны живут в жаркой и засушливой пустыне на юго-западе США, где они собирают семена, разбросанные ветром и наводнением в течение недель и месяцев.Фуражиры не используют следы феромонов; вместо этого они рассредоточиваются по ареалу кормодобывания в поисках семян [6]. Таким образом, регулирование кормодобывания в колониях муравьев-сборщиков, в отличие от медоносных пчел и видов муравьев, использующих феромоновые следы, не распределяет рабочих между пространственно фиксированными ресурсами, которые различаются по качеству и доступности [7–10]. Продолжительность похода муравьев-комбайнов за кормом может варьироваться в зависимости от наличия корма. Однако в масштабе кормового района колонии корм не истощается в течение периода кормодобывания в данный день, и жаркие и засушливые условия, а не недостаток корма, могут заставить колонии перестать кормиться.
Регулирование кормодобывающей деятельности управляет компромиссом между расходом воды и получением воды и пищи: собирающие воду теряют, находясь на улице в сухом воздухе, но колонии получают воду, метаболизируя жиры в семенах, которые они едят [11, 12]. Собирательство начинается каждое утро отдельной группой рабочих, патрульных [13, 14], которые покидают гнездо до того, как появляются фуражиры, и исследуют насыпь гнезда и область кормодобывания. Это безопасное возвращение патрульных, которое начинает поиск пищи, через встречи внутри гнезда между собирателями и возвращающимися патрульными [15].Как только начался поиск пищи, колонии муравьев-сборщиков регулируют скорость, с которой сборщики покидают гнездо, используя скорость прихода успешных сборщиков пищи, возвращающихся с пищей [16–20]. Когда муравей контактирует своими антеннами с другим муравьем, он воспринимает его кутикулярный углеводородный профиль [16]. Поскольку условия вне гнезда изменяют химический состав кутикулярных углеводородов, профили CHC зависят от задачи [21], так что в ходе контакта с усиками один муравей может определить, является ли другой собиратель.Доступный фуражир, ожидающий во входной камере внутри гнезда, побуждается покинуть гнездо за счет контакта антенн с фуражирами, несущими корм [18–20]. Скорость взаимодействия доступного фуражира внутри камеры входа в гнездо коррелирует с местной плотностью муравьев [20]. Таким образом, более высокая скорость приходящих собирателей приводит к более высокой скорости взаимодействий [19]. Поскольку каждый собиратель ищет, пока не найдет семя, скорость взаимодействия служит шумным измерением текущих условий кормодобывания [6, 22].Более высокая скорость возврата фуражей, которая отражает большее количество корма, увеличивает вероятность того, что доступные фуражи покинут гнездо в поисках корма [19, 20, 23].
В интеграционной модели [20] каждый доступный собиратель внутри гнезда собирает свидетельства от прибывающих собирателей, интегрируя свой недавний опыт контактов с усиками. Когда интегрированный стимул преодолевает порог, доступный собиратель, вероятно, покинет гнездо; в отсутствие взаимодействий собиратель, вероятно, спустится из входной камеры в более глубокое гнездо [19, 23], защищая колонию от изначально зашумленного сигнала, который возникает в результате ограниченных и неопределенных взаимодействий [24].Модель интегратора использовалась для изучения регуляции уходящей скорости кормодобывания в короткие промежутки времени, равные минутам [22].
Колонии регулируют свою деятельность по добыче пищи в более длительных временных масштабах, например, от часа к часу, от дня к дню [25, 26] и по годам [5, 25, 27, 28] по мере того, как колонии становятся старше и больше. В течение периода времени от десятков минут до часов муравьи, которые появляются в качестве доступных фуражиров внутри гнезда, покидают гнездо, чтобы добыть корм, найти семена, вернуться в гнездо и снова стать доступными фуражирами.Таким образом, активация доступных фуражиров внутри гнезда посредством взаимодействия с прибывающими фуражирами связана по «замкнутому циклу» с добычей пищи за пределами гнезда через обратную связь самих муравьев: поток муравьев-фуражиров, выходящих из гнезда, является входом для кормодобывающая деятельность, а результатом кормодобывающей деятельности является поток кормящихся муравьев в гнездо (см. рис. 1). Однако мало что известно о роли обратной связи в регулировании кормодобывающей деятельности в масштабе часов и о том, как кормодобывающая деятельность приспосабливается к изменяющимся условиям окружающей среды.К середине дня летом температура высокая, а влажность низкая (S1 Рис). Активность в поисках пищи возрастает с самого начала ранним утром, а затем стабилизируется, часто сохраняя постоянную скорость от десятков минут до часов. В жаркую полуденную жару он практически не проявляет активности.
Рис. 1. Схема модели с обратной связью с двумя компонентами внутри гнезда и одним компонентом вне гнезда.
Компонент «Взаимодействие» отображает последовательность входящих фуражиров λ в на стимул s , чтобы представить результат взаимодействий доступных фуражиров внутри камеры входа в гнездо с входящими фуражирами, несущими корм.При картировании используется негерметичный интегратор, который увеличивается на фиксированную величину с каждым приближающимся собирателем и имеет естественную скорость распада. Компонент «Отклик доступных фуражиров» отображает s на последовательность уходящих собирателей λ из с использованием нелинейной динамики осциллятора ФитцХью-Нагумо. Каждое колебание представляет собой муравей, покидающий гнездо на поиски корма. Компонент «Собирательство» преобразует λ из в λ в , используя случайную временную задержку с соответствующим распределением вероятностей, чтобы представить время, которое муравей проводит за пределами гнезда за поиском пищи.
https://doi.org/10.1371/journal.pcbi.1006200.g001
Экологическое значение имеет то, как колония регулирует добычу пищи в зависимости от условий окружающей среды, особенно температуры и влажности. Колонии живут 20-30 лет — это время жизни единственной королевы-основательницы, которая год за годом производит недолговечных рабочих. Приблизительно в возрасте пяти лет матка начинает производить репродуктивные потомства, которые спариваются с репродуктивными потомками других колоний, а дочерние королевы находят потомственные колонии [29].Колонии различаются в регулировании кормодобывания, и эти различия сохраняются из года в год, включая различия в том, как часто колонии активны [25] и в том, как они реагируют на изменение условий температуры и влажности [5, 26, 28]. Сохранение поведения при поиске пищи на протяжении многих лет в последующих когортах рабочих и сравнение родительских и потомственных колоний [29] позволяет предположить, что поведение при поиске пищи передается по наследству. Похоже, что различия между колониями в регуляции кормодобывания возникают из-за различий в том, как особи реагируют на взаимодействия с прибывающими собирателями, то есть в скорости взаимодействия, необходимой для того, чтобы стимулировать собирателя покинуть гнездо [20].
То, как колония приспосабливает добычу пищи к низкой влажности и высокой температуре, имеет решающее значение для репродуктивного успеха: колонии, которые экономят воду, с большей вероятностью будут иметь потомственные колонии [5]. Мы предполагаем, что эти различия между колониями в их реакции на различные условия окружающей среды являются результатом различий в том, как их фуражиры оценивают влажность, и в том, как это влияет на их реакцию на взаимодействия с прибывающими фуражирами [26]. Недавняя работа предполагает, что это зависит от вариаций нейрофизиологии биогенных аминов, таких как дофамин [30].Здесь мы моделируем, как колонии приспосабливаются к условиям окружающей среды, регулируя их «изменчивость», определяемую как их чувствительность к взаимодействиям с возвращающимися фуражирами. Наша цель — предложить проверяемые гипотезы об источниках различий между колониями, на которые естественный отбор может влиять на формирование коллективного поведения.
Предыдущие работы по моделированию прояснили, как исходящая скорость кормодобывания в масштабе времени в минутах зависит от входящей скорости кормодобывания [22] и как индивидуумы оценивают скорость взаимодействия [20].Но мы не знаем, как они сочетаются для корректировки кормодобывающей активности в поминутной шкале времени, как поддерживаются стабильные нормы кормодобывания, как корректировки могут зависеть от условий окружающей среды и как они могут отличаться от колонии к колонии.
Здесь мы предлагаем модель с обратной связью (рис. 1), чтобы ответить на эти вопросы, исследуя, как оценка внешних условий прибывающим собирателем обеспечивает дополнительную обратную связь с колонией и, в свою очередь, регулирует скорость сбора корма в колонии. Наша модель частично мотивирована частым использованием динамики возбудимости для моделирования нейронов, а также параллелями между взаимодействиями муравьев и муравьев, которые стимулируют добычу пищи, и взаимодействиями между нейронами, лежащими в основе когнитивных способностей организмов [20, 31–34]. ].Используя хорошо изученную динамику возбудимости слабо взаимодействующего коллектива, мы вводим обратную связь во многих временных масштабах и исследуем общие вопросы, касающиеся стабильности и отзывчивости на изменяющуюся среду.
Опираясь на теорию и инструменты динамических систем и систем управления, мы изучаем взаимосвязь в модели между быстрой активацией фуражиров внутри гнезда и медленной обратной связью от входящих фуражиров для описания с небольшим количеством параметров того, как входящие фуражиры скорость исходящего кормодобывания приспосабливается к изменяющимся условиям в масштабе времени от десятков минут до часов.Мы показываем, как стабилизируются нормы кормодобывания, и предполагаем, как небольшие различия в значениях параметров могут привести к вариациям в регулировании кормодобывания для разных условий окружающей среды и для разных колоний.
Методы
Полевые наблюдения за кормодобыванием
Мы выполнили полевые наблюдения за колониями красных муравьев-комбайнов на месте долгосрочного исследования недалеко от Родео, Нью-Мексико, США. Наблюдения проводились в августе и сентябре 2015, 2016 и 2017 гг.Собиратели покидают гнездо в ручьях или тропах, которые могут простираться на расстояние до 20 м от гнезда [35]. Каждый собиратель оставляет след в поисках семян и, найдя пищу, возвращается в гнездо [6, 35]. Данные об уровне кормодобывания регистрировались с начала периода кормодобывания с раннего утра до примерно полудня. Мы записали время, когда фуражиры пересекали линию, перпендикулярную тропе, на расстоянии около 1 м от входа в гнездо, как и в предыдущей работе (например, [22, 28, 36]). Временные метки для каждого собирателя, пересекающего линию, записывались либо вручную в режиме реального времени с помощью электронного планшета и специального программного обеспечения, либо из видеозаписей, обработанных с помощью программного обеспечения компьютерного зрения (AnTracks Computer Vision Systems, Маунтин-Вью, Калифорния).В некоторых случаях мы использовали и планшет, и видео, чтобы убедиться, что оба метода сбора данных дают одинаковые результаты.
Мы обозначаем,, последовательность раз, когда входящие фуражиры пересекают линию, и,,, последовательность раз, когда уходящие фуражиры пересекают линию. Последовательности входящих и исходящих собирателей представлены как суммы бесконечно узких идеализированных шипов в форме дельта-функций Дирака: (1) где n и m — индексы последнего входящего и выходящего собирателя, соответственно, до момента времени t .Мы оценили мгновенные входящие и исходящие скорости кормодобывания в единицах муравьев / сек, используя фильтр со скользящим окном с окном Δ t = 300 сек: (2) куда (3)
Мы выбрали размер скользящего окна, чтобы он был достаточно длинным, чтобы убрать шум, но достаточно коротким, чтобы сохранить интересные динамические характеристики скорости кормодобывания от десятков минут до часов.
Модель
Мы предлагаем низкоразмерную динамическую модель с небольшим количеством параметров, которая имеет достаточно богатую динамику, чтобы охватить диапазон наблюдаемого поведения при поиске пищи в поминутной шкале времени, и при этом сохраняет поддающуюся обработке для анализа.Мы используем модель для систематического исследования влияния параметров модели и условий окружающей среды, особенно температуры и влажности, на скорость кормодобывания.
Наша модель состоит из трех компонентов, как показано на Рис. 1: 1) Взаимодействия компонент моделирует накопление свидетельств доступными фуражирами внутри камеры входа в гнездо от их взаимодействий с приближающимися фуражирами, несущими корм, 2) Ответ доступных фуражиров компонент моделирует активацию доступных фуражиров, чтобы они покидали гнездо на корм в ответ на накопленные доказательства, и 3) компонент Foraging моделирует сбор семян за пределами гнезда активными фуражирами.Мы предполагаем, что общее количество собирателей N (активные фуражиры за пределами гнезда плюс доступные фуражиры внутри гнезда) остается постоянным в течение дня кормодобывания, хотя это предположение можно ослабить при обобщении модели.
Взаимодействия.
Мы используем динамику утечки-интегратора для моделирования стимула s , который группа доступных фуражиров внутри камеры входа в гнездо испытывает в результате их взаимодействия с прибывающими кормовыми фуражирами: (4) Интегратор утечки (4) интегрирует информацию из последовательности входящих собирателей (рис. 2A), но информация «просачивается», т.е.е., со временем медленно распадается. Здесь мы используем динамику утечки-интегратора для оценки мгновенной скорости приходящих фуражиров, которая пропорциональна общей скорости взаимодействий, испытываемых доступными фуражирами внутри гнезда [19]. Сигнал непрерывного времени s увеличивается на фиксированную величину k с каждым входящим собирателем в λ в и экспоненциально затухает обратно до нуля с постоянной времени τ (рис. 2B).
Рис 2.Модель без обратной связи.
A) Последовательность прихода фуражиров λ в . Б) Сигнал стимула с , связанный с λ в . C) FN выход v для входа s . D) Последовательность исходящих фуражиров λ из , полученная пороговым значением выхода FN снизу на 0,75.
https://doi.org/10.1371/journal.pcbi.1006200.g002
Динамика утечки интегратора работает как накопитель свидетельств, который постепенно забывает свидетельства прошлого.Эта динамика использовалась для моделирования химических синапсов [37] и использовалась в качестве модели нейронов интеграции и запуска, когда нет границы сброса [38-40].
Ответ доступных фуражиров.
Мы используем динамику возбудимости ФитцХью-Нагумо (FN) [41, 42], часто используемую для моделирования нейрональной возбудимости, в качестве феноменологической модели для активации доступных фуражиров внутри входной камеры гнезда. Наш выбор динамики мотивирован сходством между взаимодействиями муравьев с муравьями, которые активируют доступных собирателей, чтобы они покидали гнездо на корм, и взаимодействиями между нейронами, которые управляют познанием у организмов [20, 31–34].В нейрональных условиях динамика FN моделирует ответ мембранного напряжения нейрона на электрический стимул. Состояние v — это напряжение, а второе состояние u — это переменная восстановления, которая моделирует поток ионов через мембрану и обеспечивает относительно медленную отрицательную обратную связь по скорости изменения v . При низких значениях стимула напряжение остается в покое; для промежуточных значений напряжение колеблется; а при больших значениях напряжение достигает насыщения.В нашей феноменологической модели колебание представляет активацию уходящего собирателя.
Сначала мы рассматриваем однородную колонию и моделируем безразмерное скалярное состояние активации v доступных фуражиров во входной камере гнезда как переменную быстрой шкалы времени в уравнениях FN [41, 42]: (5) (6) Эти уравнения описывают динамику нелинейного осциллятора со стимулом s уравнения (4) в качестве входа и v в качестве выхода. Колебания являются результатом баланса между положительной обратной связью в v (первый член справа в уравнении (5)) и отрицательной обратной связью в безразмерной медленной шкале времени, переменной восстановления и .Параметр c , который масштабирует отрицательную обратную связь, модулирует частоту колебаний и диапазон значений стимула s , которые приводят к колебаниям. Мы вводим c в уравнения (5) и (6), так что изменение c определяет частоту колебаний и диапазон значений стимула s , которые приводят к колебаниям, но оставляет другие особенности динамика без изменений. Поскольку c регулирует отзывчивость и делает это лучше, чем параметры ϵ 1 , ϵ 2 и a , мы позволяем c представлять волатильность доступных фуражиров.
Параметр ϵ 2 определяет временное разделение между динамикой быстрого и медленного состояний, а параметр ϵ 1 определяет временное разделение между динамикой FN и динамикой стимула (4). Параметр a обеспечивает смещение к s , и его значение выбирается на основе значения k , которое представляет собой увеличение стимула s в результате взаимодействия с приближающимся собирателем.Параметр a выбирается таким образом, чтобы k было больше порога, выше которого вход s вызывает колебания. Выявление, по крайней мере, одного колебания на каждого изолированного приходящего кормораздатчика в модели позволяет быстро увеличить интенсивность кормодобывания в течение первых нескольких минут кормодобывания, когда начальная интенсивность кормодобывания низкая.
Динамика активации (уравнения (5) и (6)) доступных фуражиров дает три качественно различных динамических режима, определяемых величиной входа s и значениями бифуркации b 1 и b 2 (рис. 2С).В первом режиме система остается в состоянии покоя в течение 0 < с < b 1 . Это отражает ситуацию, в которой стимул для доступных фуражиров покидать гнездо невелик, потому что приближающихся фуражиров мало. Во втором режиме, который имеет место, когда b 1 < s < b 2 , система находится в возбужденном состоянии с колебаниями v . Это отражает ситуацию, когда приходящие фуражиры достаточно часты, чтобы стимулировать доступных фуражиров.Переход от состояния покоя к колебательному поведению при увеличении s соответствует бифуркации Хопфа, а b 1 — соответствующей точке бифуркации. Колебания проявляются в виде короткоживущих шипов, и мы определяем каждый шип, для которого v увеличивается выше 0,75, как фуражир, покидающий гнездо (Рис. 2D). Кратчайшее возможное время между выходом фуражиров из гнезда определяется изменчивостью c (см. Текст S1).
В третьем режиме, соответствующем очень большим значениям s > b 2 , колебания отсутствуют, и система фиксируется в состоянии насыщения .Переход от колебательного режима к насыщенному — это вторая бифуркация Хопфа с точкой бифуркации b 2 . Это означает, что высокая мгновенная входящая скорость, которая дает высокое значение с , приведет к насыщению в динамике FN, что приведет к снижению мгновенной исходящей скорости. Условия, представленные эффектом насыщения, включают: 1) эффекты перенаселенности, которые снижают процент взаимодействий, испытываемых каждым доступным фуражиром, по отношению к приходящей норме кормодобывания, 2) ограниченный размер входного туннеля гнезда, который ограничивает количество собирателей, которые могут войти в него, и покидать гнездо за короткий промежуток времени, и 3) разница во времени между высокими темпами исхода, в секундах, и временем, необходимым в минутах, чтобы фуражиры переместились из более глубоких камер гнезда во входную камеру [19, 23].
Собирательство.
Мы рассматриваем процесс поиска семян вне гнезда как случайную задержку по времени. Мы моделируем интервал между временем, когда собиратель покидает гнездо, и временем, когда он возвращается с пищей, как случайную величину хи-квадрат X , с параметром D , представляющим среднее время поиска пищи в минутах. Распределение времени кормодобывания F ( X , D ) составляет (7) где Γ ( X ) и Γ ( a , X ) — это гамма-функция и верхняя неполная гамма-функция, соответственно.Это распределение со смещением вправо основано на полевых наблюдениях за продолжительностью походов за пищей, измеряемой как общее время, прошедшее с момента, когда собиратель покидает гнездо, до момента, когда он возвращается с пищей [6]. Для D = 2, F ( X , 2) = 1- e — X /2 .
Наша модель процесса сбора пищи эквивалентна системе очередей [43], в которой поступающие заявки, представленные исходящими сборщиками λ из , находят начальное число после заданного случайного «времени обслуживания».Предполагается, что количество серверов в этом аналоге процесса сбора урожая как очереди бесконечно, потому что сборщикам не нужно ждать, прежде чем они начнут искать начальное число (т. Е. Перед получением услуги). В теории организации очередей очереди со случайным временем обслуживания и бесконечным числом серверов могут прояснить влияние времени обслуживания на ожидаемое количество клиентов, обслуживаемых в любое время.
Предлагаемый механизм реагирования на условия окружающей среды.
Мы предлагаем механизм реакции колонии на условия окружающей среды, проиллюстрированный на рис. 3, в котором изменчивость фуражира изменяется после того, как он совершил поход за пищей и подвергся воздействию условий за пределами гнезда.Предлагаемый механизм основан на измерениях, показывающих, что температура и влажность внутри гнезда остаются постоянными на протяжении всего периода кормодобывания (см. S1 Рис.). Это означает, что фуражиры не имеют информации об условиях на улице, пока они не покинут гнездо.
Рис. 3. Блок-схема предлагаемого механизма реакции колонии на условия окружающей среды.
Доступные фуражиры внутри гнезда состоят из двух наборов: f u соответствует тем, кто еще не покинул гнездо и поэтому не информирован об условиях за пределами гнезда, и f i соответствует информации, полученной во время предыдущего похода за пищей.Ответ каждого набора на s представлен другой моделью FN, отличающейся параметром волатильности c u для неинформированных и c i для информированных. Выходные сигналы этих двух осцилляторов взвешиваются вероятностно с использованием прореживания, чтобы получить исходящий поток собирателей λ из ( t ).
https://doi.org/10.1371/journal.pcbi.1006200.g003
В первом приближении модель изменяет изменчивость каждого фуражира после того, как он впервые покидает гнездо на корм. Доступные фуражиры, которые еще не были за пределами гнезда и поэтому не осведомлены о текущей температуре и влажности за пределами гнезда, имеют изменчивость c u . Доступные фуражиры, которые хотя бы один раз выходили на улицу для кормления и, следовательно, информированы о текущей температуре и влажности, имеют изменчивость c i .
Значения c u и c i , представляющие средний неинформированный и средний информированный доступный собиратель, могут быть любыми положительными действительными числами. Эти значения могут варьироваться в зависимости от колонии и в разные дни. Неинформированная летучесть c u может варьироваться в течение нескольких дней для данной колонии и между колониями в зависимости от потребностей колонии, таких как количество выводка, которое нужно кормить, и количество хранимой пищи, размер колонии или нейрофизиологические факторы, такие как биогенные амины (например,г. [30]). Информированная изменчивость c i отражает реакцию на условия, которые меняются как в почасовом, так и в дневном масштабе времени, такие как влажность и температура за пределами гнезда. Например, чем жарче и суше на улице, тем меньше c i , поэтому фуражиры становятся менее летучими и, следовательно, с меньшей вероятностью будут совершать последующие походы за кормом; чем прохладнее и влажнее на улице, тем крупнее c i , поэтому фуражиры становятся более летучими и, следовательно, с большей вероятностью будут совершать последующие походы за кормом. c i может варьироваться в зависимости от колонии из-за физиологических различий между колониями в ответ на условия. В настоящее время нет доказательств того, что c i зависит от того, сколько времени требуется собирателю, чтобы найти семя, но для изучения этого вопроса необходимы дальнейшие исследования.
Пусть f u будет набором n u неинформированных доступных фуражиров, которые еще не покинули гнездо в течение дня и, таким образом, не имеют информации об условиях окружающей среды, и f i набор n i проинформировал доступных фуражиров, которые подверглись воздействию условий окружающей среды во время одного или нескольких предыдущих походов за кормом в тот день.Мы предполагаем, что как только собиратель получает информацию, он остается информированным до конца дня сбора корма. Муравьи в f u имеют волатильность c u , а муравьи в f i имеют волатильность c i Пусть x u = n u / ( n u + n i ) и i / ( n u + n i ) — доля доступных фуражиров, которые не информированы и информированы, соответственно, где мы предполагаем, что n u + n i > 0.Тогда x u + x i = 1.
Первоначально x i = 0 и колония полностью неинформирована ( x u = 1). Когда фуражиры возвращаются в гнездо после первой поездки, x i начинает увеличиваться и может продолжать увеличиваться до x i = 1 ( x u = 0) , когда все фуражиры N хотя бы раз выходили за пределы гнезда.Сколько минут (или часов) требуется, чтобы x i перешел с 0 на 1, зависит от N , D и меняющейся скорости кормодобывания. Чтобы смоделировать изменяющуюся норму кормодобывания, мы используем два набора динамики осциллятора FN: один для представления реакции на s неинформированных муравьев в f u с изменчивостью c u и a второй, чтобы представить ответ на s информированных муравьев в f i с изменчивостью c i .Пусть соответствующие последовательности выходных сигналов из двух динамических осцилляторов равны λ u и λ i соответственно. Мы определяем последовательность уходящих собирателей λ из как вероятностную сумму λ u и λ i , используя метод, называемый прореживание [44]: Каждое событие на λ i сохраняется в λ out с вероятностью x i , и каждое событие в λ u сохраняется в λ out с вероятностью 1 — x я .Когда x i = 0, скорость кормодобывания определяется как c u , а при x i = 1 скорость кормодобывания определяется c i . Когда 0 < x i <1, эффективное значение c будет нелинейной комбинацией c u и c i . Чем выше эффективный c , тем выше будет выходная скорость кормодобывания.
Здесь собиратели изменяют свою изменчивость только один раз после своего первого похода за пищей на улицу. Мы обнаружили, что даже с этой корректировкой при первом воздействии модель обеспечивает наблюдаемый диапазон кормодобывающего поведения. Однако модель можно обобщить, а прогнозы уточнить, допустив корректировки в последующих походах за пищей, а также допустив другие виды корректировок. Например, более двух наборов доступных собирателей с разными значениями изменчивости можно использовать для моделирования эффектов повторяющихся воздействий на окружающую среду, изменения условий в последовательных походах или уменьшения информации о внешней среде с течением времени.Уменьшение N (общее количество фуражиров снаружи и доступных внутри гнезда) можно использовать для моделирования активных фуражиров, которые возвращаются в более глубокое гнездо после воздействия жарких и сухих внешних условий [19].
Результаты
Наблюдения за регуляцией кормодобывания рыжих муравьев-комбайнов
Наблюдения за мгновенными темпами кормодобывания, рассчитанными по данным 2015, 2016 и 2017 годов, показывают, что по колониям и дням приходящие и исходящие коэффициенты кормодобывания r в ( t ) и r из ( t ), где t — время суток, претерпевают переходный процесс (т.е.е., временный образец изменения) в начале периода кормодобывания с последующим уравновешиванием до почти равного значения, т. е. r в ( t ) ≈ r из ( т ), в середине кормодобывающего периода.
Уравновешивание приходящей и исходящей норм кормодобывания до почти равного значения длится в течение интервалов от десятков минут до нескольких часов, и поэтому мы называем это квазистационарным режимом (QSS).Мы показываем данные для двух колоний на рис. 4. Мы наносим на график входящую скорость r в (синий) и исходящую скорость r из (красный), вычисленную по данным для колонии 1357 ( Рис. 4A) и Колония 1317 (Рис. 4B) в зависимости от времени суток 20 августа 2016 г. Для Колонии 1357 показатели уравновешивались до почти равного значения в начале дня, то есть между 8:00 и 8:30 утра. За этим последовала пара динамических корректировок, но затем к 9:30 утра до самого полудня, когда все муравьи вернулись в гнездо, входящие и исходящие скорости были очень близко уравновешены при скорости QSS около 0.25 муравьев / сек. Колония 1317 также достигла QSS. Его входящие и исходящие скорости уравновесились до почти одинакового значения вскоре после 10:00, что продолжалось до самого полудня, когда все муравьи вернулись в гнездо. Колонии сильно различаются по скорости кормодобывания [28], и это верно и для этих двух. Для колонии 1317 скорость QSS составляла приблизительно 0,65 муравьев / сек, что более чем вдвое превышало скорость QSS для колонии 1357 в тот же день.
Рис. 4. Графики входящей скорости кормодобывания r в (синий) и исходящей скорости кормодобывания r из (красный) в зависимости от времени суток 20 августа 2016 г. для A) Колония 1357 и Б) Колония 1317.
Квазистационарное состояние (QSS), при котором входящие и исходящие скорости уравновешиваются до почти равного значения, может наблюдаться для обеих колоний. Показатель QSS для колонии 1317 был более чем в два раза выше, чем для колонии 1357.
https://doi.org/10.1371/journal.pcbi.1006200.g004
Мы показываем данные для двух других колоний на рис. 5. Рис. 5A и 5C показывают r в (синий) и r из (красный) по сравнению с временем суток для одной колонии, Колония 664, в два разных дня: 27 августа 2015 г. и 31 августа 2015 г.На каждом графике можно увидеть, что ставки приходят к почти одинаковому значению где-то после 10:30 утра. Мы наносим зеленый цвет на график кумулятивной разницы между количеством входящих и исходящих собирателей в зависимости от времени суток. Скорости соответствуют QSS, когда зеленая кривая приблизительно горизонтальна. Эти данные показывают, как было замечено ранее [45], что скорость кормодобывания в данной колонии меняется день ото дня, демонстрируя, что кормление регулируется процессами, отличными от количества собирателей в колонии, которое остается относительно постоянным во временной шкале. месяцев.Из рис. 5A и 5C видно, что 27 августа 2015 года Colony 664 достигла уровня QSS, который более чем вдвое превышает уровень QSS 27 августа 2015 года. Мы отмечаем, что 27 августа 2015 года было немного прохладнее и влажнее. чем 31 августа 2015 года. 27 августа средняя температура и влажность составляли 25,3 C и 58%, а 31 августа — 26,8 C и 53%. Более того, в 11:00 27 августа они были 27,5 ° C и 52%, а в 11:00 31 августа — 28,8 ° C и 45%. На рис. 5E показаны данные для колонии 863 на 1 сентября 2015 г., записанные вручную.QSS не наблюдается, т. Е. Муравьи вышли, но затем вернулись в гнездо к 11:00, не поддерживая стабильное состояние фуражиров вне гнезда. Колония 863 действительно достигла QSS при достаточно высокой скорости кормодобывания в 11:00 5 сентября 2015 г. (см. Рис. S2A). Эти наблюдения согласуются с измерениями, показывающими, что 1 сентября 2015 года было намного жарче и суше, чем 5 сентября 2015 года. 1 сентября средняя температура и влажность составляли 25,2 C и 53%, а 5 сентября — 22.6 C и 77%. См. Более подробную информацию в таблице S1 SI.
Рис. 5. Графики данных по скорости кормодобывания.
Графики временных рядов показывают входящий коэффициент кормодобывания r в (синий), исходящий коэффициент кормодобывания r выход (красный) и совокупную разницу между количеством входящих и исходящих собирателей (зеленый) по сравнению со временем суток. Графики ввода-вывода показывают r out ( t ) по сравнению с r в ( t ) с цветовой шкалой, представляющей время дня t .A) и B) Колония 664 27 августа 2015 г. C) и D) Колония 664 31 августа 2015 г. E) и F) Колония 863 1 сентября 2015 г.
https://doi.org/10.1371/journal.pcbi.1006200.g005
Рисунки 4 и 5 представляют наблюдения, которые предполагают, что уравновешивание приходящей и исходящей норм кормодобывания до почти равной скорости является важной характеристикой регулирование кормодобывания в колониях красных муравьев-комбайнов. Кроме того, уравновешенная скорость и возможность раннего прекращения кормодобывания зависят от факторов, которые различаются между колониями (рис. 4) и ото дня к дню (рис. 5).Мы исследуем переходные процессы в нормах кормодобывания далее на рис. 5. В начале дня кормодобывания как r в , так и r из быстро увеличивались с r из увеличивались быстрее. чем r в . Это привело к быстрому увеличению количества активных фуражиров вне гнезда. За быстрым увеличением обеих скоростей последовало снижение r из до уравновешенного почти равного значения QSS (рис. 5A и 5C) или раннее возвращение муравьев в гнездо (рис. 5E). ).
Графики ввода-вывода показывают соотношение между входящими и исходящими темпами кормодобывания Рис. 5B, 5D и 5F. На этих рисунках показаны те же данные, что и на рис. 5A, 5C и 5E, соответственно, но график r out ( t ) против r in ( t ) со временем суток t в часах, обозначенных цветовой шкалой. Переходные нормы в начале дня кормодобывания выглядят как изогнутые траектории над диагональю, потому что r out ( t ) обычно выше, чем r in ( t ). ).На рис. 5B и 5D кривая поднимается, а затем опускается до значения QSS, при котором траектория затем уравновешивается вокруг точки на диагонали, соответствующей равным входящим и исходящим скоростям. Этот подъем и спад кривой на графике ввода-вывода типичен, даже когда траектория возвращается в исходное положение, как в случае на рис. 5F.
Данные, показанные на рисунках 4 и 5, а также на рис. S2, представляют данные, собранные в 2015, 2016 и 2017 гг. Температура и влажность для этих наборов данных приведены в таблице S1.S2B На рис. Показан еще один пример очень раннего прекращения кормодобывания. S2C и S2D Рис. Показывает два разных примера длительных переходных процессов. S2E и S2F На рис. Показаны два примера резкого увеличения количества уходящих кормов в начале кормодобывающего дня. См. Подробности в тексте S2.
Динамика модели
Динамика кормодобывания внутри гнезда.
Учитывая последовательность входящих муравьев λ в , наша модель динамики поиска пищи внутри гнезда без обратной связи (рис. 1) предсказывает соответствующую последовательность выходящих муравьев λ из .Мы находим аналитическое приближение для сопоставления средней поступающей нормы кормодобывания со средней уходящей скоростью кормодобывания, параметризованной изменчивостью c . Для этого мы предполагаем, что λ в является пуассоновским процессом с (постоянной) средней скоростью поступления; это оправдано для наблюдений за входящими и уходящими последовательностями фуражиров в течение коротких периодов времени [22].
Присваиваем значениям параметров модели k = 0,3, τ = 0,41, a = 0.35, ϵ 1 = 0,2 и ϵ 2 = 0,05, что обеспечивает богатое динамическое поведение. В то время как качественное поведение не меняется для различных значений ϵ 2 ≪ 1, очень высокие или низкие значения a , k и / или τ дают динамику, в которой стимул s либо слишком низкий или слишком высокий, чтобы вызывать колебания. Таким образом, значения для a , k и τ выбраны для уравновешивания их противоположных эффектов на s и колеблющуюся область FN.
Осциллирующая область динамики FN соответствует диапазону значений s между точками бифуркации FN b 1 и b 2 , вычисляемым как. Смещение a не влияет на размер колеблющейся области, тогда как волатильность c может управлять им: по мере увеличения c размер колеблющейся области уменьшается (S3 рис.). Смещение a изменяет нижнее пороговое значение s , т.е.е., нижняя точка бифуркации b 1 , выше которой колеблется ФН. Чтобы гарантировать, что каждый изолированный входящий собиратель вызывает по крайней мере одного выходящего фуражира, учитывая k , что является увеличением s для взаимодействия с входящим собирателем, мы выбираем a так, чтобы k > b 1 для всех c ∈ [0, 5]. Мы выбираем k и τ , чтобы получить разумные значения s для диапазона приходящей кормовой активности, наблюдаемой в данных.Одна из сильных сторон модели заключается в том, что, хотя есть некоторая гибкость в выборе , , k, и τ, , чтобы они удовлетворяли этим желаемым условиям, на качественное поведение системы конкретные значения не влияют. выбрано.
Для очень низкого уровня является низким, потому что s низкий, и система FN остается в состоянии покоя с редкими короткими периодами колебательного поведения (рис. 2). Для очень высокого значения также является низким, поскольку с является высоким, и система FN большую часть времени остается в состоянии насыщения.Напротив, высокий для этого дает s , который удерживает систему FN внутри колеблющейся области. В колеблющейся области равна частоте колебаний, которая обратно пропорциональна волатильности c , как показано в S1 Text.
Чтобы получить выражение для собственной частоты колебаний в FN, мы вычисляем приближение для его периода T LC ( s , c ), которое использует временное разделение между динамиками из v и u , см. текст S1 и рис. S4.При допущении входящей скорости Пуассона процесс s является эргодическим (см. S3 Text). Таким образом, в течение достаточно длительных периодов времени подходящая статистика по времени сходится к статистике ансамбля, что позволяет нам аппроксимировать долю времени, которую с проводит в колеблющейся области, используя функцию плотности вероятности с в установившемся состоянии. Мы вычисляем в S4 Text как кусочную функцию, где кусочные элементы удовлетворяют рекуррентным уравнениям и зависят от k и τ .Исходя из этого, мы можем построить аналитическое выражение для как функции от c (см. S3 Text): (8)
На рис. 6A мы построили график зависимости от использования уравнения (8) для различных значений c . Полученные в результате кривые ввода-вывода разомкнутого контура, которые мы называем вложенными кривыми ввода-вывода , показывают, что аналитическое отображение из в в нелинейно зависит от c . Увеличивающаяся крутизна кривой на низком уровне становится более выраженным для более высокого c , потому что частота колебаний пропорциональна c .Точно так же уменьшение крутизны кривых для высоких значений также становится более выраженным для высоких значений c . Это связано с тем, что по мере увеличения c b 2 уменьшается, вызывая насыщение FN при более низких значениях. Максимальное значение имеет место при s , что удерживает систему FN внутри колеблющейся области. Из-за этого максимум должен быть меньше или равен собственной частоте колебаний при заданном значении c .
Рис. 6.
A) Аналитические приближения для кривых ввода / вывода гнезда. B) Смоделированные кривые ввода-вывода гнезда для различных значений c . Каждая пара планок погрешностей соответствует 10 имитационным испытаниям продолжительностью каждые 5 минут с постоянной ожидаемой скоростью поступления и постоянной волатильностью c . Пунктирная черная линия представляет точки, в которых средняя входящая скорость равна средней исходящей скорости.
https://doi.org/10.1371/journal.pcbi.1006200.g006
На рис. 6B показаны кривые ввода-вывода, полученные при моделировании разомкнутой системы для различных постоянных входных скоростей Пуассона при фиксированной волатильности. В каждом случае мы измерили итоговую среднюю исходящую скорость. Мы установили λ в на пятиминутный процесс Пуассона, и в каждом из 10 испытаний моделирования мы записали выходной сигнал λ из . Мы вычислили среднюю скорость исходящего кормодобывания, разделив общее количество исходящих собирателей в испытании на 300 секунд, в течение которых продолжалось испытание.Мы использовали среднее значение 10 испытаний в качестве точечной оценки как функцию данного параметра волатильности c . Мы построили гнездовые кривые ввода-вывода, повторив этот процесс оценки точек для двенадцати различных значений, сохраняя постоянное значение c .
Смоделированные кривые ввода-вывода на рис. 6B хорошо согласуются с аналитическими кривыми ввода-вывода на рис. 6A. Кривые моделирования немного выше, чем аналитические кривые, потому что T LC недооценивает период колебаний FN (см. S4 Рис.) И потому, что уравнение (8) полагается на то, что s является эргодическим (см. S3 Текст).Тот факт, что существует хорошее соответствие между кривыми моделирования, вычисленными из 5-минутных входных последовательностей, и аналитическими кривыми, полученными в предположении бесконечного периода времени, предполагает, что временная статистика с ( t ) быстро сходится к ансамблю статистика. Эта быстрая сходимость указывает на то, что наше аналитическое приближение справедливо и для коротких временных масштабов. Мы используем это в нашем анализе динамики модели с обратной связью.
Точки, в которых кривые ввода-вывода гнезд на рис. 6 пересекаются с черной пунктирной диагональной линией, соответствуют (квази) установившимся решениям при одинаковой входящей и исходящей скорости кормодобывания, наблюдаемой в данных.Рис. 6 предполагает, что для достаточно высоких значений c , равная интенсивность кормодобывания положительна и отделена от нуля, фиксируя нетривиальную установившуюся норму кормодобывания, как на рис. 4 и 5B и 5D. Тем не менее, Рис. 6 предполагает, что для низких значений c , равная скорость кормодобывания близка к нулю, фиксируя установившееся состояние с незначительным кормлением, как на Рис. 5F.
Чтобы понять, как c влияет на равную норму кормодобывания, примите во внимание, что, поскольку k > b 1 , каждый изолированный входящий собиратель вызывает по крайней мере одно колебание в выходе FN и поэтому при низком уровне равен или больше, чем .При высоких значениях c частота колебаний в FN высока, и изолированный входящий собиратель вызовет более одного колебания, в результате чего кривые ввода-вывода гнезда с начальным наклоном больше единицы и пересечением с диагональной линией на единственная точка от начала координат. Напротив, для низких значений c изолированный входящий собиратель вызовет ровно одно колебание, в результате чего кривые ввода-вывода гнезда с начальным наклоном равны единице, т. Е. Кривая лежит на диагональной линии близко к началу координат и нигде не пересекается. еще.
Этот анализ предполагает наличие критического значения c *, такого, что если c > c *, существует изолированное стационарное решение вдали от источника, и если c < c *, стационарное решение остается близким к началу координат. Мы находим такую оценку сверху, которая достаточна для существования изолированного стационарного решения вдали от начала координат. Для b 1 < k < b 2 можно показать, что количество колебаний, вызванных одним входящим фуражиром, не превышает (- τ log b 1 / к ) / T LC .Мы численно решили это выражение для c , используя асимптотическое разложение T LC , описанное в S1 Text, и обнаружили, что FN колеблется по крайней мере два раза на каждого приходящего собирателя. Следовательно, это достаточное условие для того, чтобы кривая ввода-вывода гнезда лежала выше диагональной линии на низком уровне и пересекала диагональную линию в изолированной точке, что соответствует нетривиальному установившемуся уровню кормодобывания.
Динамика кормодобывания вне гнезда.
Учитывая последовательность исходящих собирателей λ на выходе со скоростью r на выходе , динамика кормодобывания вне гнезда предсказывает соответствующую отложенную последовательность приходящих собирателей λ в со скоростью r в . Мы используем результаты теории массового обслуживания, чтобы найти аналитические выражения, связывающие r из до r в и ожидаемое количество активных фуражиров вне гнезда.
Для облегчения анализа мы предполагаем, что λ из является неоднородным пуассоновским процессом (т.е. пуассоновским процессом с изменяющейся во времени скоростью) [22]. Применяя известные результаты для очередей с неоднородным пуассоновским распределением времен прихода [46], получаем следующие три результата:
- Пусть Q ( t ) представляет количество активных фуражиров вне гнезда, тогда для каждого раза t ′ = t /60, где t — секунды, Q ( t ′) имеет распределение Пуассона со средним значением (9)
- Выходной процесс, описывающий, как фуражиры покидают систему очередей, то есть процесс λ в , описывающий, как фуражиры возвращаются в гнездо, является неоднородным пуассоновским процессом со средним (10)
- r in относится к r out by (11)
Уравнения (7) и (9) показывают, как количество активных фуражиров вне гнезда зависит от истории уходящих фуражиров.Уравнение (10) показывает, что если исходящий процесс поиска пищи является неоднородным процессом Пуассона, то входящий процесс сбора пищи также является неоднородным процессом Пуассона. А уравнение (11) показывает, как интенсивность поступающего кормодобывания r в зависит от истории исходящего коэффициента кормодобывания r из .
В установившемся режиме скорость исходящего кормодобывания постоянна, т. Е. r out ( t ) = r *, а уравнение (11) сокращается до r in = r из = r *, т.е.е., поступающая скорость кормодобывания также постоянна и равна исходящей норме кормодобывания. Более того, уравнение (9) сводится к (12) то есть среднее количество активных собирателей за пределами гнезда рассчитывается как установившаяся скорость кормодобывания r *, умноженная на среднее время похода за кормом D .
Время релаксации для процесса вывода очереди для достижения установившегося состояния может быть проанализировано с учетом скорости поступления ступенчатой функции r out ( t ′) = 0 для t ′ <0 и r out ( t ′) = r * для t ′ ≥ 0.Тогда из уравнения (11) для t ′ ≥ 0. Разница между скоростью ввода и вывода очереди как функция времени равна (13) для t ′ ≥ 0. Для иллюстрации, мы вычисляем для D = 2, что правая часть уравнения (13) упрощается до r * e — t ′ / 2 и Очередь нагула экспоненциально сходится во времени к установившемуся состоянию, когда входная и выходная скорости равны.
Замкнутая модель динамики.
В нашей модели уходящие фуражиры из гнезда выходят за кормом, возвращаются в гнездо как входящие фуражиры после нахождения семени, а затем снова выходят за фуражом, если они достаточно возбуждены (рис. 1).Здесь мы показываем, что добавление обратной связи от исходящих к прибывающим фуражирам к динамике разомкнутого контура в гнезде дает долгосрочную динамику со стабильным и притягивающим равновесием, при котором входящие и исходящие коэффициенты равны. Стабильность изолированного равновесия подразумевает устойчивость: уравновешенная в установившемся состоянии скорость кормодобывания сохраняется даже при наличии возмущений, например, небольших изменений в норме приходящих собирателей. Когда изменчивость c > c *, критическое значение, определенное ранее, установившаяся интенсивность кормодобывания нетривиальна, тогда как если c < c *, установившаяся интенсивность кормодобывания незначительна.
Для динамики внутри гнезда мы показали, что c параметризует семейство кривых ввода-вывода гнезда, описываемых уравнением (8), которые отображаются в коротких временных масштабах в несколько минут. Для c < c * кривая ввода / вывода всегда имеет наклон меньше или равный 1, так что она лежит на или ниже диагональной линии, где r in = r из . Для c > c * кривая ввода / вывода вложения имеет начальный наклон больше 1, а затем пересекает диагональную линию r in = r out в нетривиальной точке.Для динамики вне гнезда мы показали, что отображение r out ( t ) на r in ( t ) описывается в более длительных временных масштабах, составляющих десятки минут. задержкой по времени, задаваемой уравнением (11).
Мы изучаем динамику модели с обратной связью для шкал времени от десятков минут до часов, исследуя поведение дискретного итерационного отображения r n = G c ( r n −1 ), где r n представляет собой среднюю норму кормодобывания на временном шаге n и r n −1 представляет собой среднюю скорость кормодобывания на временном шаге n — 1 .Мы можем интерпретировать r n и r n −1 как среднюю приходящую норму или среднюю исходящую норму кормодобывания, поскольку средняя входящая частота становится средней исходящей частотой после временной задержки. Отображение определяется зависимыми от c кривыми ввода-вывода гнезда, показанными на рис. 6. G c описывает процесс, посредством которого поступающая норма кормодобывания становится скоростью исходящей корма через динамику активации собирателей внутри. гнездо, которое затем становится поступающей нормой кормодобывания в более позднее время.
Когда G c лежит ниже (выше) диагональной линии, где r in = r out , среднее количество исходящих фуражиров на каждого входящего фуражира меньше ( больше), чем один, и итерации G c уменьшаются (увеличиваются) r (рис. 7). Для c > c *, G c имеет одну нестабильную фиксированную точку в начале координат и одну привлекательную стабильную фиксированную точку, где r дюйм = r out .Для c < c *, G c имеет небольшой интервал фиксированных точек, близких к началу координат. Таким образом, динамика модели с замкнутым контуром эволюционирует во времени либо в сторону конечной установившейся скорости кормодобывания r in = r out = r * (Рис.7, c = 2 и c = 5) или незначительным кормлением (рис.7, c = 0,1).
Рис. 7. Динамика модели, иллюстрирующая реакцию норм кормодобывания на условия окружающей среды.
Красные, фиолетовые и синие кривые показывают траектории замкнутого контура r out ( t ) по сравнению с r in ( t ) для фиксированной волатильности c , равной 5.0, 2.0 и 0.1 соответственно. Изначально все доступные собиратели не осведомлены об окружающей среде и обладают изменчивостью c u = 5,0. Темно-серая пунктирная кривая показывает динамику в случае, когда фуражиры, находящиеся в окружающей среде, снижают свою изменчивость до c i = 2.0, что может случиться в умеренно жаркий и сухой день. Светло-серая пунктирная кривая показывает динамику в случае, когда фуражиры, находящиеся в окружающей среде, снижают свою изменчивость до c i = 0,1, как это может случиться в очень жаркий и засушливый день.
https://doi.org/10.1371/journal.pcbi.1006200.g007
Стабильность уравновешенной нормы кормодобывания в стационарном состоянии и последствия для устойчивости являются результатом баланса между положительной обратной связью от входящих муравьев, активирующих большее количество исходящих муравьев. муравьи и отрицательная обратная связь от эффектов насыщения.Величина и дисперсия установившейся нормы кормодобывания увеличиваются с c . Величина также зависит от k и τ , поскольку они влияют на s в уравнении (8), которое можно численно решить, чтобы найти, как величина изменяется с c (рис. 8A). Мы называем устойчиво устойчивую норму кормодобывания QSS.
Рис. 8.
A. Аналитическая величина квазистационарной (QSS) скорости кормодобывания, полученная путем численного решения уравнения (8).Б. Моделирование модели с обратной связью для 7 различных значений волатильности c . Первоначальная последовательность входящих собирателей для всех симуляций была установлена равной последовательности входящих собирателей, зарегистрированной в течение первых 11 минут для Колонии 859 20 августа 2017 г., которая имеет среднюю скорость прихода 0,01 муравьев / сек. Общее время для всех симуляций составило 3 часа. Среднее время кормодобывания составляло 10 минут ( D = 10).
https://doi.org/10.1371/journal.pcbi.1006200.g008
Как показано на рис. 8B, моделирование модели с обратной связью подтверждает предсказания итерированной модели отображения (рис. 7). Мы инициализируем динамику кормодобывания, задав λ в от t = 0 до t = 60 × ( D + 1) секунд, чтобы оно было равно начальной последовательности прибывающих собирателей для Колонии 859 20 августа. , 2017, который имеет очень низкую среднюю скорость прихода 0,01 муравьев / сек в течение первых 15 минут (S2C, рис.). Используя дополнительную минуту, т.е.е., D + 1 вместо D , для начальной последовательности входящих фуражиров помогает гарантировать, что последовательность входящих фуражиров не закончится внезапно до того, как первые несколько уходящих фуражиров вернутся в гнездо.
Динамика с обратной связью в зависимости от условий окружающей среды.
Для пуассоновских последовательностей входящих собирателей средняя скорость исходящего кормодобывания колонии дается как взвешенная сумма выходов неинформированных и информированных: (14)
Динамику замкнутого цикла все еще можно изучать как итеративное отображение, но мы позволяем отображению развиваться во времени, G c = G c ( t ), от начальное сопоставление с изменчивостью c u до окончательного сопоставления с изменчивостью c i .Темные и светло-серые кривые на рис. 7 иллюстрируют, как карта G c ( t ) изменяется со временем, когда c u = 5,0 и c i = 2,0 или c i = 0,1. Сначала динамика развивается вдоль (красный), но по мере увеличения x i динамика все больше смещается к, а траектория на графике r из ( t ) против r в ( t ) перемещается к кривой c i .В случае c i = 2,0, траектория сходится к фиксированной точке, связанной с c = 2,0 (более темная серая пунктирная кривая). В случае c i = 0,1 траектория сходится к единственной фиксированной точке, которая является исходной точкой, что приводит к прекращению кормодобывания (светло-серая пунктирная кривая).
На рис. 9 показаны результирующие графики временных рядов и входов-выходов для трех различных симуляций модели с механизмом реакции на условия окружающей среды.Моделирование различается набором из четырех параметров: c u , c i , N и D . Смоделированные траектории качественно напоминают траектории полевых наблюдений, показанные на рис. 5. Мы устанавливаем начальную последовательность прибывающих собирателей, как на рис. 8B.
Рис. 9. Моделирование замкнутой модели с механизмом адаптации.
Графики имеют ту же форму, что и на рис. 5, и можно проводить качественные сравнения между A и B здесь и рис. 5A и 5B, между C и D здесь и рис. 5C и 5D, и между E и F здесь и рис. 5E. и 5F.A) и B) c u = 3, c i = 0,9, N = 500, D = 5. C) и D) c u = 3, c i = 0,75, N = 200, D = 5. E) и F) c u = 5, c i = 0,02, N = 600, D = 15.
https: // doi.org / 10.1371 / journal.pcbi.1006200.g009
На рис. 9A и 9B показаны результаты для c u = 3, c i = 0,9, N = 500 и D = 5. В этом случае c u намного выше, чем c i , что приводит к системе с перерегулированием, в которой исходящая скорость кормодобывания увеличивается быстрее, чем входящая скорость, а затем уменьшается, прежде чем установится около устойчивого состояния, когда ставки примерно равны 0.7 муравьев / сек. Это качественно похоже на наблюдения колонии 664 27 августа 2015 г. на рис. 5A и 5B. Результатом относительно небольшого общего количества собирателей N и короткого среднего времени кормодобывания D является то, что доля информированных собирателей быстро увеличивается, что приводит к быстрой конвергенции к установившемуся состоянию. Чистое количество собирателей вне гнезда в установившемся состоянии колеблется с небольшой изменчивостью и составляет около 230, что близко к предсказанию, даваемому уравнением (12).
На рис. 9C и 9D показаны результаты для c u = 3, c i = 0.75, N = 200 и D = 5. Этот случай моделирует ту же колонию, что и на рис. 9A и 9B, но в более жаркий и сухой день, когда общее количество муравьев N , занятых добычей пищи, может быть уменьшена, и изменчивость информированных муравьев c i может быть уменьшена. За перерегулированием следует, что скорость кормодобывания стабилизируется примерно на уровне 0,25 муравьев / сек. Результатом низких значений N и D является то, что доля осведомленных фуражиров увеличивается очень быстро, что приводит к быстрой конвергенции к устойчивому состоянию.Это качественно похоже на наблюдения колонии 664 31 августа 2015 г. на рис. 5C и 5D.
На рис. 9E и 9F показаны результаты моделирования для c u = 5, c i = 0,02, N = 600, D = 15. В этом случае c i близка к нулю, что приводит к тому, что колония уходит на корм, но затем возвращается в гнездо без продолжительного кормления. Результатом длительного среднего времени кормодобывания D является то, что доля осведомленных собирателей увеличивается медленными темпами, что приводит к более длительной переходной динамике к установившемуся состоянию.Это качественно похоже на наблюдения колонии 664 27 августа 2015 г. на рис. 5E и 5F.
Время, необходимое для перехода колонии от полностью неинформированной к полностью информированной о внешних условиях, определяется условиями c u , c i , D , N и начальные условия для r в и r из . Низкие значения c u приводят к изначально низкой скорости исходящего кормодобывания, так что соответствующая скорость, с которой фуражиры становятся информированными, тоже низкая, даже если c i является высокой (S5A Рис.).Низкие значения c i могут вызвать длительные переходные процессы, потому что, как только критическое количество собирателей становится информированным, низкая изменчивость затрудняет информирование оставшихся собирателей. Высокие значения D и N также могут приводить к длительным переходным процессам, поскольку время, необходимое для перехода в полностью информированное состояние, зависит от количества доступных фуражиров и от того, сколько времени требуется информированным сборщикам, чтобы вернуться в гнездо (S5B Рис).Наконец, изначально высокие значения r из приводят к быстрому увеличению числа активных фуражиров, сокращая время, необходимое для достижения информированного состояния с темпами кормодобывания, которые достигают QSS (S5C Рис). Качественные сравнения могут быть выполнены между моделированием в S5A, S5B и S5C Fig, и данными в S2C, S2D, S2E и S2F Fig, соответственно.
Обсуждение
Мы вывели и проанализировали низкоразмерную аналитическую модель динамики кормодобывания, которая требует лишь небольшого числа параметров, чтобы качественно охватить широкий диапазон переходных и устойчивых характеристик, наблюдаемых в нормах кормодобывания колоний красных муравьев-комбайнов.Наша модель расширяет предыдущую работу, используя обратную связь в нескольких временных масштабах, чтобы учесть, как скорость кормления в гнездо и из гнезда изменяется в течение длительного времени, от десятков минут до часов.
Важно отметить, что большие временные рамки позволяют провести на основе модели исследование того, как колония без централизованного контроля и небольшого количества индивидуальной информации о состоянии колонии или окружающей среды может стабильно регулировать свою норму кормодобывания и реагировать на температуру и влажность за пределами вложенность в поминутную шкалу времени.Стабильность подразумевает устойчивость установившейся нормы кормодобывания к небольшим возмущениям, например, небольшим изменениям в норме приходящих собирателей. Кроме того, поскольку модель поддается анализу, ее можно использовать для систематического получения эмпирически проверяемых прогнозов кормодобывающего поведения как функции критических параметров модели, включая количество собирателей N , среднее время похода за кормом D и изменчивость c . В нашей модели эти параметры определяют постоянную норму кормодобывания, независимо от начальной нормы кормодобывания.Однако переходный процесс и время сходимости к установившемуся режиму зависят от начальных скоростей; более высокие начальные скорости приводят к более быстрой сходимости. Модель предполагает, что изменение летучести, когда фуражиры подвергаются воздействию температуры и влажности за пределами гнезда, может объяснить наблюдаемое поведение при поиске пищи в различных условиях окружающей среды. Кроме того, модель предполагает, что различия между колониями в летучести в ответ на температуру и влажность могут вызывать наблюдаемые различия между колониями в регулировании кормодобывания.
Наша модель и анализ подчеркивают важность обратной связи в различных временных масштабах в регулировании кормодобывающей деятельности. Предыдущая работа изолирует динамику разомкнутого цикла внутри гнезда, которая отображает входящих и исходящих муравьев в очень короткие сроки. Мы обращаемся к поминутной шкале времени, аналитически исследуя динамику замкнутого цикла, которая связывает медленную добычу пищи за пределами гнезда с быстрой активацией фуражиров внутри гнезда через обратную связь, генерируемую самими муравьями и их взаимодействием с другими.Поток кормящихся муравьев из гнезда является входом для кормодобывающей деятельности, а вывод кормовой активности — это поток кормовых муравьев в гнездо. Эффективная изменчивость колонии также изменяется в модели в масштабе времени от минут до часов, поскольку фуражиры впервые покидают гнездо и подвергаются воздействию внешней влажности и температуры, что обеспечивает гибкость в регулировании кормодобывающей деятельности от минуты до минуты. -часовые сроки.
В модели изменчивость c аппроксимирует среднюю чувствительность имеющихся фуражиров в гнезде к взаимодействиям с входящими фуражирами: чем выше c , тем меньше взаимодействий необходимо для активации доступных фуражиров для выхода и кормления.Связь между c и активацией собирателей нелинейна, и тонкости нашей модели отражают некоторые сложности системы. Мы используем аналитические прогнозы, чтобы показать, как c определяет три важных характеристики динамики модели кормодобывания (см. Рис. 8): 1) начальный переходный процесс в нормах кормодобывания входящих и исходящих, параметризованных с помощью c , 2) уравновешивание входящих и исходящих кормлений. исходящие нормы кормодобывания до стабильной и, следовательно, устойчивой, квазистационарной нормы, параметризованной c , и 3) прогнозом раннего прекращения кормодобывания или отказа от кормодобывания вообще, если c < c *, a значение критической волатильности c *.
Поведение разных колоний в один и тот же день или одной и той же колонии в разные дни соответствует в модели различным значениям c . Более низкие значения c приводят к динамике модели, которая согласуется с данными для более жарких и засушливых дней, потому что более низкие значения c означают, что доступные фуражиры менее изменчивы и, следовательно, с меньшей вероятностью будут выходить за кормом. Более высокие значения c приводят к динамике модели, которая согласуется с данными для более прохладных и более влажных дней, потому что более высокие значения c означают, что доступные фуражиры более изменчивы и, следовательно, с большей вероятностью будут выходить за кормом.Модель отличает изменчивость c u доступных фуражиров в гнезде, которые еще не отправились в поход за пищей, от изменчивости c i доступных фуражиров в гнезде, которые уже были вне гнезда и подвергались воздействию окружающей среды (рис. 3). Результатом является переход от кормодобывающей активности муравьев с изменчивостью c u к поисковой активности муравьев с изменчивостью c i , которая может длиться от нескольких минут до нескольких часов. N муравьи выходят в другое время во время своего первого похода за кормом и возвращаются в гнездо после кормления в среднем на D минут (рис. 7).
Различия между колониями в значениях параметров c u , c i , N и D могут привести к наблюдаемым нами различиям в поведении при поиске пищи между колониями. Действительно, в диапазоне значений четырех параметров c u , c i , N и D модель описывает диапазон переходных и квазистационарных состояний. Динамика скорости кормодобывания наблюдалась в данных, собранных для колоний красных сборщиц в августе и сентябре 2015, 2016 и 2017 годов.Таким образом, модель предлагает гипотезы о физиологических процессах, которые могут привести к различным значениям параметров, например, различия между колониями в том, как уходящие фуражиры реагируют на взаимодействия с возвращающимися фуражирами, и различия между колониями в том, как фуражиры реагируют на такие условия, как влажность.
Модель представляет случай, когда фуражиры корректируют свою изменчивость только после своего первого похода за кормом. Чтобы учесть большую изменчивость в пределах колонии, модель может быть обобщена на M > 2 группы доступных фуражиров в гнезде, различающиеся M значениями летучести c 1 ,…, c M .Например, обобщение можно использовать для учета собирателей, которые вносят коррективы в свою реакцию на взаимодействия в гнезде после последующих походов за кормом из-за многократного воздействия или изменения температуры и влажности. Обобщение можно также использовать для учета разложения информации о тех фуражирах, которые остаются в гнезде в течение длительного периода после похода за пищей, или для представления фуражиров, которые возвращаются в более глубокое гнездо после воздействия жарких и сухих внешних условий.
Модели кормодобывания, которые рассматривают регуляцию кормодобывающей деятельности, как правило, делятся на две категории: многоагентные модели, отслеживающие каждую особь [7–9], и компартментные модели, отслеживающие временную эволюцию фракций особей, занятых конкретная задача [10, 47].Многоагентные модели позволяют детально моделировать динамику кормодобывания, часто полагаясь на симуляции из-за их сложности и плохой аналитической управляемости. В отличие от этого, компартментные модели во многих случаях обеспечивают высокую управляемость, но предполагают очень большие размеры групп, что влияет на прогнозы, когда размер группы невелик. Наша модель рассматривает два идеализированных процесса, активацию фуражиров внутри гнезда и сбор семян вне гнезда, чтобы создать динамическую систему с небольшим количеством уравнений и параметров.Модель подходит для групп любого размера и сохраняет достаточную управляемость для создания прогнозов воздействия критических параметров модели.
Наша биологически информированная, низкоразмерная и просто параметризованная модель позволяет систематически исследовать механизмы и чувствительность, которые могут объяснить коллективное поведение и направлять дальнейшие теоретические и экспериментальные исследования. Наше использование хорошо изученной динамики возбудимости открывает путь для сравнения с другими сложными системами, такими как нейронные сети, которые управляются возбудимой динамикой.Модель вместе с нашим анализом, основанным на теории динамики и управления, способствует лучшему пониманию роли обратной связи в различных временных масштабах в коллективном поведении.
Дополнительная информация
S1 Рис. Влажность и температура на поверхности пустынной почвы и внутри камеры входа в гнездо.
Показания влажности и температуры, зарегистрированные на поверхности пустынной почвы (синий цвет) и внутри камеры входа в гнездо (красный цвет). Датчики температуры и влажности ibutton были размещены снаружи, но близко ко входу в гнездо на пустынной почве, незатененными, а внутри гнезда в вырытой яме, которая была обнаружена при раскопках, а затем закрыта сверху стеклом и затемнена.Влажность и температура снаружи гнезда значительно менялись в утренние часы, в то время как влажность и температура внутри камеры у входа в гнездо оставались относительно постоянными. Измеренное умеренное повышение температуры внутри гнезда, вероятно, связано с тем, что свет проникает во входную камеру гнезда через стекло. А) Влажность 29 августа 2014 г. (Колония E). Б) Температура 29 августа 2014 г. (Колония E). В) Влажность 31 августа 2015 г. (Колония 10). Г) Температура 31 августа 2015 г. (Колония 10).Д) Влажность 1 сентября 2015 г. (Колония 10). Е) Температура 1 сентября 2015 г. (Колония 10).
https://doi.org/10.1371/journal.pcbi.1006200.s001
(TIF)
S2 Рис. Дополнительные полевые наблюдения за скоростью кормодобывания.
Входящий коэффициент кормодобывания r в (синий), исходящий коэффициент кормодобывания r из (красный), а также разница между количеством входящих и исходящих собирателей (зеленый) в зависимости от времени суток.A) Колония 863 5 сентября 2015 г. достигла высокого уровня QSS; сравните с рис. 4E, когда в более жаркий и сухой день, 1 сентября 2015 г., колония 863 рано вернулась в гнездо. Б) Колония D19 08.08.2016 рано вернулась в гнездо; день был очень жарким и сухим. В) Колония 859 20 августа 2017 г .; переходный процесс начался поздно утром. День был прохладным и влажным. Г) Колония 1107 16 августа 2017 г .; переходный процесс был медленным. День выдался засушливый. Д) Колония 1017 23 августа 2016 г .; начальный переходный процесс был больше похож на наплыв уходящих собирателей.День выдался засушливый. Е) Колония 1015 18 августа 2016 г .; еще один начальный всплеск уходящих собирателей. День был очень сухим.
https://doi.org/10.1371/journal.pcbi.1006200.s002
(TIF)
S3 Рис. Функция плотности вероятности для функции стимула.
Каждая кривая представляет PDF p функции стимула s для различных значений входящей скорости r в . Серые прямоугольники представляют размер колебательной области в системе FN ( b 1 , b 2 ) для a = 0.35 и разные значения волатильности c . Для всех кривых k = 0,3, τ = 0,41.
https://doi.org/10.1371/journal.pcbi.1006200.s003
(TIF)
S5 Рис. Дополнительные модели замкнутой системы с механизмом адаптации.
Графики напоминают наблюдаемое поведение кормодобывания на рис. S2. Можно провести качественные сравнения между A здесь и рис. S2C, между B здесь и S2D рис. И между C здесь и на S2E и S2F рис.A) c u = 0,9, c i = 2,2, N = 500, D = 5. Настройка c u <
c i , где c u близко к c *, приводит к долгому периоду, прежде чем ставки возрастут. B) c u = 1, c i = 1, N = 1000, D = 15.Установка большого среднего времени похода за кормом D на большое приводит к длительным переходным процессам. C) c u = 0,7, c i = 0,9, N = 1000, D = 7. Установка начального λ в равным последовательности из первые 5 минут λ в для Колонии 1017 23 августа 2016 г. дают поведение, показанное на S2E и S2F Рис., которое следует за начальным всплеском фуражиров.https://doi.org/10.1371/journal.pcbi.1006200.s005
(TIF)
S1 Таблица. Температура и относительная влажность в Родео, Нью-Мексико.
Средняя температура, средняя относительная влажность, температура в 11 часов утра и относительная влажность в 11 часов утра в Родео, Нью-Мексико, США, в течение нескольких дней с данными, нанесенными на рисунки 3 и 4 и рисунок S2. Данные, собранные станцией программы Citizen Weather Observer Program E8703 и доступ через Weather Underground. Станция расположена в 2,7 км от места исследования.
https://doi.org/10.1371/journal.pcbi.1006200.s006
(PDF)
Благодарности
Мы благодарим многих преданных своему делу полевых ассистентов бакалавриата за их помощь: Сэма Кроу, Элеонору Глокнер, Кристофера Джексона, Сару Цзян, Га-Иль Ли, Артура Местаса, Бекку Нельсон и особенно Ребиа Хан.
Список литературы
- 1. Гордон Д.М. Организация работы в социальных колониях насекомых. Природа. 1996. 380: 121–124.
- 2. Самптер DJ.Коллективное поведение животных. Принстон: Издательство Принстонского университета; 2010.
- 3. Леонард NE. Мультиагентная системная динамика: бифуркация и поведение групп животных. Annu Rev Control. 2014. 38 (2): 171–183.
- 4. Хопфилд Дж. Нейронные сети и физические системы с возникающими коллективными вычислительными возможностями. Proc Natl Acad Sci USA. 1982. 79 (8): 2554–2558. pmid: 6953413
- 5. Гордон Д.М. Награды за сдержанность в коллективном регулировании кормодобывания колониями муравьев-комбайнов.Природа. 2013; 498 (7452): 91–93. pmid: 23676676
- 6. Беверли Б.Д., МакЛендон Х., Наку С., Холмс С., Гордон Д.М. Как верность местоположения приводит к индивидуальным различиям в кормовой активности муравьев-сборщиков. Behav Ecol. 2009. 20 (3): 633–638.
- 7. Тениус Р., Шмикль Т., Крайльсхайм К. Оптимизация энергетики пчелиной семьи с помощью социального обучения на основе задержек в очереди. Связь науки. 2008. 20 (2-3): 193–210.
- 8. Кокс MD, Myerscough MR.Гибкая модель кормодобывания колонией медоносных пчел: влияние индивидуального поведения на успех кормления. Журнал теоретической биологии. 2003. 223 (2): 179–197. pmid: 12814601
- 9. Schmickl T, Crailsheim K. Затраты на колебания окружающей среды и преимущества динамических децентрализованных решений о кормлении медоносных пчел. Адаптивное поведение. 2004; 12 (3-4): 263–277.
- 10. Грановский Б., Латти Т., Дункан М., Самптер Д. Дж., Бикман М. Как танцующие медоносные пчелы следят за изменениями: роль пчел-инспекторов.Поведенческая экология. 2012. 23 (3): 588–596.
- 11. Лайтон Дж., Варфоломей Г.А. Стандартный энергетический метаболизм пустынного муравья-сборщика Pogonomyrmex rugosus: влияние температуры, массы тела, размера группы и влажности. Proc Natl Acad Sci USA. 1988. 85 (13): 4765–4769. pmid: 16593953
- 12. Lighton JR, Feener DH Jr. Скорость потери воды и проницаемость кутикулы у собирателей пустынного муравья Pogonomyrmex rugosus. Physiol Zool. 1989. 62 (6): 1232–1256.
- 13.Гордон Д.М. Динамика переключения задач у муравьев-комбайнов. Поведение животных. 1989. 38 (2): 194–204.
- 14. Гордон Д.М. Поведенческая гибкость и экология кормодобывания муравьев-семенников. Американский натуралист. 1991. 138 (2): 379–411.
- 15. Грин MJ, Гордон DM. Кутикулярные углеводороды информируют о решениях задач. Природа. 2003 05; 423: 32 EP -. pmid: 12721617
- 16. Грин MJ, Гордон DM. Социальные насекомые: кутикулярные углеводороды информируют о решениях задач.Природа. 2003. 423 (6935): 32–32. pmid: 12721617
- 17. Грин MJ, Гордон DM. Скорость взаимодействия информирует комбайн о решениях задач. Behav Ecol. 2007. 18 (2): 451–455.
- 18. Грин MJ, Пинтер-Уоллман N, Гордон DM. Взаимодействие с комбинированными химическими сигналами сообщает муравьям-сборщикам о решении покинуть гнездо в поисках пищи. PLoS One. 2013 01; 8 (1): e52219. pmid: 23308106
- 19. Пинтер-Уоллман Н., Бала А., Меррелл А., Кейроло Дж., Стамп М.К., Холмс С. и др.Муравьи-сборщики используют взаимодействия, чтобы регулировать активацию и доступность собирателей. Anim Behav. 2013. 86 (1): 197–207. pmid: 24031094
- 20. Дэвидсон Д.Д., Арауко-Алиага Р.П., Ворон С., Гордон Д.М., Голдман М.С. Влияние взаимодействий между муравьями-жнецами на решения собирателей. Передняя часть Ecol Evol. 2016; 4: 115. pmid: 28758093
- 21. Вагнер Д., Тиссот М., Гордон Д. Рабочая среда изменяет углеводородный состав кутикулы муравьев-комбайнов. J Chem Ecol. 2001. 27 (9): 1805–1819.pmid: 11545372
- 22. Прабхакар Б., Дектар К.Н., Гордон Д.М. Регулирование кормовой активности колонии муравьев без пространственной информации. PLoS Comput Biol. 2012; 8 (8): e1002670. pmid: 22927811
- 23. Плесс Э., Кейроло Дж., Пинтер-Уоллман Н., Кроу С., Аллен К., Матур М.Б. и др. Взаимодействия увеличивают доступность и активность собирателей у муравьев-сборщиков. PloS One. 2015; 10 (11): e0141971. pmid: 26539724
- 24. Файнерман О., Корман А. Индивидуальное и коллективное познание у социальных насекомых.J Exp Biol. 2017; 220 (1): 73–82. pmid: 28057830
- 25. Гордон Д.М. Поведенческая гибкость и экология кормодобывания муравьев-семеедов. Am Nat. 1991; с. 379–411.
- 26. Гордон Д.М., Дектар К.Н., Пинтер-Уоллман Н. Изменчивость колоний муравьев-комбайнов в кормодобывающей активности и реакции на влажность. PLoS One. 2013; 8 (5): e63363. pmid: 23717415
- 27. Гордон Д.М. Как рост колонии влияет на вторжение фуражиров между соседними колониями муравьев-комбайнов.Behav Ecol and Sociobiol. 1992. 31 (6): 417–427.
- 28. Гордон Д.М., Гетц А., Грин М.Дж., Холмс С. Колонические вариации в коллективном регулировании кормодобывания муравьями-комбайнами. Behav Ecol. 2011. 22 (2): 429–435. pmid: 22479133
- 29. Гордон Д.М. Развитие ареала кормления муравьиной колонии. Anim Behav. 1995. 49 (3): 649–659.
- 30. Фридман Д.А., Пилько А., Сковронска-Кравчик Д., Красинская К., Паркер Дж. В., Хирш Дж. И др. Роль дофамина в коллективной регуляции кормодобывания у муравьев-хищников.iScience. 2018; 8: 283–294. pmid: 30270022
- 31. Пассино К.М., Сили Т.Д., Вишер П.К. Познание роя у медоносных пчел. Behav Ecol and Sociobiol. 2008. 62 (3): 401–414.
- 32. Couzin ID. Коллективное познание в группах животных. Trends Cogn Sci. 2009. 13 (1): 36–43. pmid: 192
- 33. Маршалл Дж. А., Богач Р., Дорнхаус А., Планке Р., Ковач Т., Франкс Н. Р.. Об оптимальном принятии решений в мозгах и колониях социальных насекомых. Интерфейс J R Soc. 2009. 6 (40): 1065–1074.pmid: 19324679
- 34. Маршалл Дж. А., Фрэнкс Н. Р.. Познание на уровне колонии. Curr Biol. 2009; 19 (10): R395 – R396. pmid: 19467202
- 35. Гордон Д.М. Развитие ареала кормления муравьиной колонии. Anim Behav. 1995. 49 (3): 649–659.
- 36. Гордон Д.М., Холмс С., Наку С. Кратковременное регулирование кормодобывания у муравьев-комбайнов. Behav Ecol. 2008. 19 (1): 217–222.
- 37. Рот А., Ван Россум MC. Моделирование синапсов. В: Шуттер Э.Д., редактор.Методы компьютерного моделирования для нейробиологов. Кембридж: Пресса Массачусетского технологического института; 2009. с. 139–160.
- 38. Лапик Л. Речерчес количественно оценивает электрическое возбуждение с характерной поляризацией. J Physiol Pathol Gen.1907; 9 (1): 620–635.
- 39. Таквелл Х. Введение в теоретическую нейробиологию. Vol. 1. Линейная теория кабеля, дендритная структура и стохастические теории. Кембриджские исследования по математической биологии. Кембридж: Издательство Кембриджского университета; 1988 г.
- 40. Штейн РБ. Теоретический анализ нейрональной изменчивости. Биофиз Дж. 1965; 5 (2): 173. pmid: 14268952
- 41. ФитцХью Р. Импульсы и физиологические состояния в теоретических моделях нервной мембраны. Биофиз Дж. 1961; 1 (6): 445. pmid: 19431309
- 42. Нагумо Дж., Аримото С., Йошизава С. Активная линия передачи импульсов, имитирующая аксон нерва. Proc IEEE Inst Electr Electron Eng. 1962. 50 (10): 2061–2070.
- 43. Купер РБ.Введение в теорию массового обслуживания. 2-е изд. Вашингтон, округ Колумбия: Северная Голландия; 1981.
- 44. Льюис П.А., Шедлер Г.С. Моделирование неоднородных пуассоновских процессов утонением. Нав Рес Логист. 1979. 26 (3): 403–413.
- 45. Гордон Д.М., Холмс С., Наку С. Кратковременное регулирование кормодобывания у муравьев-комбайнов. Behav Ecol. 2008. 19 (1): 217–222.
- 46. Эйк С.Г., Мэсси В.А., Уитт В. Физика очереди Mt / G / ∞. Oper Res. 1993. 41 (4): 731–742.
- 47.Самптер Д., Пратт С. Модель моделирования для понимания поиска пищи социальными насекомыми. Поведенческая экология и социобиология. 2003 февраль; 53 (3): 131–144.
Сравнительная структура сообществ муравьев-хищников в засушливых районах Австралии и Северной Америки по JSTOR
АбстрактныйВ засушливой зоне Австралии видовое богатство муравьев больше, а видов грейфоровых млекопитающих меньше, чем в пустынях Северной Америки. Это исследование было направлено на то, чтобы определить, отличается ли структура сообществ муравьев-собирателей от таковой в пустынях Северной Америки, уделяя особое внимание различиям, связанным с малочисленностью грызунов.Мы проверили три гипотезы: (1) поскольку грызунов меньше, австралийские муравьи-хищники должны быть более многочисленными и разнообразными в местных средах обитания, чем в пустынях Северной Америки с аналогичной продуктивностью; (2) поскольку отсутствие грызунов позволит муравьям использовать более крупные семена, которые предпочитают грызуны в Северной Америке, австралийские сообщества муравьев должны включать больший диапазон размеров и содержать более крупных рабочих; и (3) что помимо различий, вызванных малочисленностью грызунов, австралийские и североамериканские сообщества будут сближаться по характеристикам структуры сообществ в результате конкуренции.Мы отобрали образцы 19 сообществ через климатический градиент в засушливой зоне Австралии и сравнили результаты с данными, полученными ранее для 10 сообществ Северной Америки. Австралийские муравьи-собиратели демонстрировали аналогичное альфа-разнообразие (внутри среды обитания), но более высокое бета-разнообразие (между средами обитания) между сообществами. Сообщества Австралии и Северной Америки были схожи по видовому богатству, видовому разнообразию, количеству общих видов и численности муравьев, хотя австралийские сообщества, как правило, были богаче и разнообразнее в местах с меньшим количеством осадков.В североамериканских сообществах увеличилось видовое богатство, разнообразие, количество обычных видов и пропорции видов, питающихся колоннами, с увеличением количества осадков. Напротив, австралийские сообщества не менялись регулярно по градиенту осадков. Таким образом, первая гипотеза была отвергнута. Вторая гипотеза также была отвергнута, потому что австралийские муравьи-собиратели были меньше, охватывали меньший диапазон размеров и имели тенденцию более плотно распределяться по градиенту размеров. Данные о питании указывают на несоответствие между использованием ресурсов и их доступностью, предполагая, что австралийские общины, возможно, обычно не находятся в равновесии со своей ресурсной средой.Однако ни одна из первых двух гипотез не может быть проверена однозначно, поскольку остается возможным, что эволюционное взаимодействие между семенами и муравьями в отсутствие грызунов позволило семенам принять защитные механизмы, сводящие к минимуму потребление муравьями, или что другие зерноядные (особенно птицы) частично компенсируют это. за малочисленность грызунов. Проверка третьей гипотезы была неоднозначной. Некоторые результаты предполагают основные сходства между эффектами конкуренции на сообщества, например, одинаковые максимальные значения численности, богатства и видового разнообразия.Однако некоторые признаки австралийских сообществ отличались от таковых в Северной Америке: добыча пищи происходила в более широком диапазоне температур почвы; заметным было временное смещение кормодобывания среди сосуществующих видов; и отдельные виды, добывающие пищу, встречались с одинаковой частотой в зависимости от климатических градиентов. Мы обсуждаем различные биотические и абиотические особенности, которые могут объяснить различия в структуре сообществ между континентами. Мы пришли к выводу, что малочисленность грызунов, потенциально конкурирующей группы зерноядных животных, не привела к предсказуемым изменениям в сообществах муравьев-собирателей засушливой Австралии.
Информация о журналеВидение экологических монографий заключается в том, что они должны стать местом для публикации интегративных, синтетических статей, которые разрабатывают новые направления в области экологии. В этом мире стремительного научного прогресса и нескончаемых изменений окружающей среды должно быть место для продуманной интеграции научных идей, данных и концепций, которые питают разум и направляют развитие развивающейся науки об экологии.«Экологические монографии» предоставляют эту комнату с обширным обзором устойчивого будущего.
Информация об издателеWiley — глобальный поставщик решений для работы с контентом и контентом в областях научных, технических, медицинских и научных исследований; профессиональное развитие; и образование. Наши основные направления деятельности производят научные, технические, медицинские и научные журналы, справочники, книги, услуги баз данных и рекламу; профессиональные книги, продукты по подписке, услуги по сертификации и обучению и онлайн-приложения; образовательный контент и услуги, включая интегрированные онлайн-ресурсы для преподавания и обучения для студентов и аспирантов, а также для учащихся на протяжении всей жизни.Основанная в 1807 году компания John Wiley & Sons, Inc. уже более 200 лет является ценным источником информации и понимания, помогая людям во всем мире удовлетворять их потребности и реализовывать их чаяния. Wiley опубликовал работы более 450 лауреатов Нобелевской премии во всех категориях: литература, экономика, физиология и медицина, физика, химия и мир. Wiley поддерживает партнерские отношения со многими ведущими мировыми сообществами и ежегодно издает более 1500 рецензируемых журналов и более 1500 новых книг в печатном виде и в Интернете, а также базы данных, основные справочные материалы и лабораторные протоколы по предметам STMS.Благодаря растущему предложению открытого доступа, Wiley стремится к максимально широкому распространению и доступу к публикуемому контенту, а также поддерживает все устойчивые модели доступа. Наша онлайн-платформа, Wiley Online Library (wileyonlinelibrary.com), является одной из самых обширных в мире междисциплинарных коллекций онлайн-ресурсов, охватывающих жизнь, здоровье, социальные и физические науки и гуманитарные науки.
Агентство по охране окружающей среды США, Этикетка для пестицидов, малатион 851 г / л + гамма-цигалотрин 12.8 г / л ЭК, 23.02.2015
% PDF-1.6 % 1 0 объект > поток application / pdf
Задние бета- и передние гамма-колебания предсказывают когнитивную проницательность | Журнал когнитивной неврологии
Наше ЭЭГ-исследование нейронной основы инсайта и трансформирующего мышления во время решения проблем дало результаты, которые сохраняются в широком диапазоне условий и сравнений.
Как и предполагалось, наблюдалось снижение мощности бета-диапазона (13–30 Гц) в теменной, теменно-затылочной и центрально-височной областях мозга, когда человек преимущественно занимался преобразующим рассуждением.Когнитивная обработка данных иногда связана с изменением бета-мощности (Pesonen et al., 2007; Bastiaansen et al., 2005; Weiss & Rappelsberger, 1998). Кроме того, снижение бета-мощности в теменной и теменно-затылочной областях наблюдается в сочетании с независимым от модальности увеличением нагрузки на память (Pesonen et al., 2006, 2007), которая, как полагают, необходима для трансформирующего мышления. Таким образом, наши результаты убедительно указывают на важную роль бета-колебаний в задачах трансформирующего мышления, которые критически задействуют семантическую память.
Повышенная мощность в гамма-диапазоне (30–70 Гц) и повышенное вовлечение правого полушария головного мозга (правая лобная, лобно-центральная и центрально-височная области) также наблюдались, когда участники правильно решали проблему понимания и когда они сообщали о том, что испытывают «Ага!» Правая префронтальная кора головного мозга активируется при проблемах, требующих трансформации смены установок (Goel & Vartanian, 2005; Miller & Tippett, 1996), которая представляет собой тип реструктуризации, а также при выполнении задач, связанных с последовательным мышлением, когда конфликт логики и убеждений вызывает изменение. в процессе рассуждений (Goel, Buchel, Frith, & Dolan, 2000).Кроме того, правое полушарие, по-видимому, помогает сделать доступным набор альтернативных и менее вероятных значений слов в лексической задаче (Coney & Evans, 2000), что соответствует созданию множества решений в задачах понимания. Любопытно, что мощность гамма-излучения была ниже при успешных (хитовых) испытаниях, чем при неудачных испытаниях, в период времени сразу после представления подсказки. Одна из версий состоит в том, что подсказка ограничила пространство поиска до такой степени, что период пост-подсказки не требовал истинных преобразующих рассуждений при некоторых испытаниях.Сравнение пост-намеков было единственным из четырех сравнений, которые не включали момент ответа, и альтернативная версия заключается в том, что усиленная гамма-мощность может быть нейронным коррелятом момента субъективного понимания, но не реструктуризации и других процессов трансформирующего мышления, которые предшествовать этому. Более того, различия в пространственном расположении увеличения (FC6, F8) и уменьшения (CP6, C4) мощности гамма-излучения предполагают наличие двух различных систем гамма-отклика. Для решения этих проблем необходимы дополнительные эксперименты.Тем не менее, на основе прошлых и настоящих открытий, мы можем обоснованно утверждать, что правая префронтальная область и колебания гамма-диапазона участвуют в трансформирующих рассуждениях.
Критически важно, что настоящие результаты подтверждают нашу четвертую и последнюю гипотезу, а именно, что изменение осцилляторной активности, лежащее в основе трансформирующего мышления, представляет собой бессознательные когнитивные , а не аффективные процессы: в отличие от того факта, что аффективное «Ага!» процессы обычно происходят примерно в момент реакции (Jung-Beeman et al., 2004), изменения в спектральной мощности, соответствующие трансформирующему мышлению в нашем исследовании, произошли за несколько секунд за до ответа . Примеры включают повышенную активность гамма-диапазона для растворов HIT по сравнению с растворами FP в правой лобно-центральной области мозга за 8 → 1 секунду до ответа (рис. 3A), снижение активности бета-диапазона в испытаниях nH_HIT по сравнению с H_HIT за 6 секунд до ответа (рис. 2C). ), и усиление верхней альфа-активности (13,5 Гц) в правой теменно-затылочной области за 7 секунд до представления подсказки по испытаниям SH по сравнению с UH (фактор 3 на дополнительном рисунке 3).Проницательные специалисты, решающие проблемы, обычно не могут сообщить о внутренних психических процессах, предшествующих их проницательному решению (Ohlsson, 1992), или радоваться, прежде чем решить возникшую проблему; следовательно, изменения в колебательных ритмах, которые происходят задолго до сознательной реакции, вряд ли будут соответствовать ощущениям радости или облегчения, которые сопровождают решение проблемы, а скорее соответствуют подсознательным (и, возможно, автоматическим) мысленным рекомбинациям очевидно далеких, но отдаленно связанных репрезентаций до того, как раствор (Simonton, 1999; Seifert et al., 1995; Metcalfe & Wiebe, 1987).
Наши результаты также подтверждают предположение о том, что предшествующее состояние мозга, а именно уровень психической подготовленности, способствует более поздней реструктуризации представления проблемы: разница в активности мозга была еще до того, как была дана подсказка между головоломками, которые в конечном итоге были решены. и те, которых не было. В отличие от Kounios et al. (2008), никаких существенных различий за период времени до представления проблемы не наблюдалось; различия наблюдались в течение периода времени после представления проблемы, но до предъявления подсказки.Это эффективно доказывает существование состояния мозга, которое восприимчиво к включению новой информации (подсказки) с существующим когнитивным представлением.
До сих пор мы сосредоточились на общих особенностях колебательных паттернов мозговой активности в четырех сравнениях. Ясно, что есть также шаблоны, которые уникальны для каждого (дополнительные рисунки 2–5). Например, активность гамма-диапазона в правой теменно-затылочной области выше при «Ага!» против «не-Ага!» решения (дополнительный рисунок 5).Этот результат совместим с сообщениями об эмоционально нагруженных стимулах, усиливающих активность гамма-диапазона в правой теменной области (Афтанас, Рева, Варламов, Павлов, & Махнев, 2003; Кейл и др., 2001), аналогичные данные, отделяющие аффективные от когнитивных процессов в контекст исследования нейронных коррелятов юмора (Goel & Dolan, 2001), и с нашей точки зрения, что рассуждения, ведущие к пониманию и результирующему «Ага!» не имеют идентичных нервных субстратов.
Сделаем несколько критических замечаний.Сначала наши участники двигали рукой, нажимая кнопку после решения задачи. Бета ERD часто описывается как нейронный коррелят для подготовки и выполнения движений; следовательно, можно утверждать, что наши основные открытия в отношении бета-ERD соответствуют подготовительной двигательной активности, а не трансформирующим рассуждениям. Однако оба условия каждого попарного сравнения либо завершались одинаковым нажатием кнопки (HIT против FP, nH_HIT против H_HIT, HR против LR), либо были отмечены общим событием (т.е., намек в SH vs.UH). Таким образом, подготовительная двигательная активность была либо обычной в данном сравнении, либо отсутствовала вообще. Кроме того, подготовительная двигательная активность, или потенциал готовности, как его иногда называют, в основном представлена частотами ниже 3 Гц (Libet, Gleason, Wright, & Pearl, 1983; Kornhuber & Deecke, 1965), тогда как наши эффекты были в основном в более высокие частоты бета (21–30 Гц) и гамма (> 30 Гц). В общем, бета-ERD не может соответствовать двигательной подготовке. Во-вторых, наш выбор меры анализа требует пояснения.Один из способов анализа многомерных данных, таких как наш, — рассматривать каждый канал, частоту и момент времени изолированно и статистически измерять надежность попарной разницы в сигнале в каждой точке этого многомерного пространства отдельно. Контролируя множественные сравнения, обычно получают отдельные точки во времени, пространстве и частоте, которые значительно различаются между двумя тестируемыми условиями. Учитывая, что не существует априорной гипотезы, столь специфичной во времени, частоте или пространстве, разрешение измерения как таковое не так точно, а моменты времени, частоты и особенно электроды не являются изолированными, независимыми единицами, это больше разумно вместо этого думать о нейронном корреляте трансформирующего мышления в терминах кластеров в многомерном пространстве.PARAFAC — это, по сути, инструмент исследования данных (Morup, Hansen, & Arnfred, 2007), сила которого заключается в выявлении регионов, где можно найти различия в пространстве, времени и частоте между двумя состояниями. Необработанная статистика F была введена в анализ PARAFAC для определения кластеров статистически значимых регионов в этом пространстве. Результаты анализа PARAFAC необходимо подтвердить с помощью необработанных частотно-временных графиков, что мы и сделали (рисунки 1, 2 и 3). Для перекрестной проверки наших результатов PARAFAC мы повторно проанализировали наши необработанные данные СКР по изолированным единицам частоты и пространства с помощью многомерного статистического теста (Hemmelmann, Horn, Süsse, Vollandt, & Weiss, 2005), основанного на частоте ложных открытий (Benjamini & Hochberg, 1995), который исправляет множественные сравнения; опять же, эффекты бета-диапазона оказались наиболее заметной статистически значимой характеристикой трансформирующего мышления среди четырех сравнений.В общем, PARAFAC — полезный, мощный способ изолировать значительную часть данных ЭЭГ большой размерности. В-третьих, у ЭЭГ плохое пространственное разрешение; результаты, локализующие функцию в отдельных областях мозга, следует интерпретировать с осторожностью. Более того, участие подкорковых структур мозга, таких как гиппокамп, в решении задач инсайта (Luo & Niki, 2003) недооценивается в ЭЭГ. В-четвертых, каждое попарное сравнение в чем-то уникально. Следовательно, нужно быть осторожным, прежде чем утверждать, что бета- и гамма-осцилляции являются нервным субстратом понимания.С другой стороны, можно с полным основанием утверждать, что эффекты более устойчивы и устойчивы в этих частотных диапазонах. Наконец, наши головоломки различаются по содержанию; Тем не менее, все они имеют один общий элемент: каждый требует трансформирующего мышления.
В заключение, наши результаты показывают, что инсайт — это отчетливый спектральный, пространственный и временной паттерн бессознательной нейронной активности, соответствующий когнитивным процессам до принятия решения, а не самооценке своего инсайта или эмоциональному «Ага!» что сопровождает решение проблемы.Будущие исследования определят общность наших результатов для других классов проблем и то, как модуляция паттернов нейронной активности с помощью умных поведенческих манипуляций или манипуляций с компьютерным интерфейсом мозга влияет на проницательность.
% PDF-1.5 % 1 0 объект > / OCGs [8 0 R] >> / Страницы 2 0 R / Тип / Каталог >> эндобдж 34 0 объект > поток 2021-08-02T01: 48: 29-07: 002006-09-29T12: 15: 30-07: 002021-08-02T01: 48: 29-07: 00uuid: 36d39cbf-bdcf-46bb-8e76-2f9ef05729d1uuid: 7084053c- 1dd2-11b2-0a00-880098c59cffapplication / pdf конечный поток эндобдж 2 0 obj > эндобдж 29 0 объект > / Ресурсы> / Шрифт> / T1_1> / T1_2> / T1_3> / T1_4 38 0 R >> / ProcSet [/ PDF / Text / ImageB] / Properties> / XObject >>> / Type / Page >> эндобдж 23 0 объект > / Ресурсы> / Шрифт> / T1_1> / T1_2> / T1_3> / T1_4> / T1_5> / T1_6 38 0 R >> / ProcSet [/ PDF / Text / ImageB / ImageC] / Свойства> / XObject >>> / Тип / страница >> эндобдж 18 0 объект > / Resources> / Font> / T1_1> / T1_2> / T1_3> / T1_4> / T1_5 38 0 R >> / ProcSet [/ PDF / Text / ImageB] / Properties> / XObject >>> / Type / Page >> эндобдж 13 0 объект > / Resources> / Font> / T1_1> / T1_2> / T1_3> / T1_4> / T1_5 38 0 R >> / ProcSet [/ PDF / Text / ImageB] / Properties> / XObject >>> / Type / Page >> эндобдж 5 0 obj > / Ресурсы> / Шрифт> / T1_1> / T1_2> / T1_3 38 0 R >> / ProcSet [/ PDF / Text / ImageB] / Properties> / XObject >>> / Type / Page >> эндобдж 35 0 объект > / ProcSet [/ PDF / Text / ImageC] / XObject >>> / Type / Page >> эндобдж 47 0 объект [54 0 R 55 0 R 56 0 R 57 0 R 58 0 R] эндобдж 48 0 объект > поток q 540.0594177 0 0 68.6011963 35.9702911 675.3988037 см / Im0 Do Q BT / T1_0 1 Тс 10 0 0 10 85,56995 558,99982 тм (1988; 48: 1740-1744.) Tj / T1_1 1 Тс -5.55699 0 Тд (Рак Res \ 240) Tj / T1_0 1 Тс 0 1 ТД (\ 240) Tj 0 1.00001 TD (Грегори Т. Спир, Донна М. Полнок, Дайан О. Хелгесон и др.) / T1_2 1 Тс 0 1 ТД (\ 240) Tj / T1_3 1 Тс 18 0 0 18 30 598,99994 тм (Интерлейкин-1 клеток HL-60) Tj 12,50396 1 тд (для индукции и секреции) Tj -0,33333 Ц -0,556 0 Тд (3) Tj 0 Ц -11.94796 0 Тд (1,25-дигидроксивитамин D) Tj 27.58085 1 тд (и) Tj / T1_4 1 Тс -0.41099 0 Тд (g) Tj / T1_3 1 Тс -27.16986 0 Td (Требование дифференциальных сигналов обоих интерферонов-) Tj ET 30 504 552 35 рэ 0 0 мес. S BT / T1_0 1 Тс 11 0 0 11 120.94202 511.99997 тм (\ 240) Tj / T1_3 1 Тс -7,55696 1 тд (Обновленная версия) Tj ET BT / T1_2 1 Тс 10 0 0 10 141 503,99994 тм (\ 240) Tj / T1_0 1 Тс 22,62295 1 тд () Tj 0 0 1 рг -22.62295 0 Тд (http://cancerres.aacrjournals.org/content/48/7/1740)Tj 0 г 0 1.00001 TD (Последнюю версию этой статьи можно найти по адресу:) Tj ET BT / T1_2 1 Тс 10 0 0 10 30 483.99997 Тм (\ 240) Tj 0 1 ТД (\ 240) Tj ET BT / T1_2 1 Тс 10 0 0 10 30 463,99997 тм (\ 240) Tj Т * (\ 240) Tj ET BT / T1_2 1 Тс 10 0 0 10 30 443,99997 тм (\ 240) Tj Т * (\ 240) Tj ET 30 329 552 115 рэ 0 0 мес. S BT / T1_0 1 Тс 11 0 0 11 120.94202 411.99997 тм (\ 240) Tj / T1_3 1 Тс -5.66901 1 тд (Оповещения по электронной почте) Tj ET BT / T1_0 1 Тс 10 0 0 10 295,4996 424 тм (относится к этой статье или журналу.) Tj 0 0 1 рг -15.44996 0 Тд (Зарегистрируйтесь, чтобы получать бесплатные уведомления по электронной почте) Tj ET BT 0 г / T1_0 1 Тс 11 0 0 11 120,94202 378,99994 тм (\ 240) Tj / T1_3 1 Тс -6.38997 1 тд (Подписки) Tj 0,556 1,00001 тд (Отпечатки и) Tj ET BT / T1_0 1 Тс 10 0 0 10 141 381,99994 тм (\ 240) Tj 13,46496 1 тд (.) Tj 0 0 1 рг -6.85098 0 Тд ([email protected]) Tj 0 г -6.61398 0 Тд (Отделение) Tj 0 1.00001 TD (Чтобы заказать перепечатку статьи или подписаться на журнал, свяжитесь с нами \ t Публикации AACR) Tj ET BT / T1_0 1 Тс 11 0 0 11 120.94202 356.99997 тм (\ 240) Tj / T1_3 1 Тс -5. \ q
Тонкая красота: Lancia Gamma Coupe
1979 годаLancia Gamma 1979 года выпуска — прекрасный пример изящно красивого купе в стиле Pininfarina, показанного здесь на 2.5 литров, форма 140 л.с. Благодаря интересному четырехцилиндровому двигателю SOHC с питанием от карбюратора (на ряд) мощность передается на передние колеса через 5-ступенчатую механическую коробку передач, и, по словам продавца, этот конкретный пример является одним из всего 6689 автомобилей. Они также заявляют о двух предыдущих частных владельцах, и автомобиль по-прежнему находится в Германии, где он был впервые продан. Кажется, что он находится в прекрасной форме, и звуки также остаются без изменений. Его можно найти здесь, в магазине Classic Cars Dornstetten в Дорнштеттене, Германия, за 9900 евро (около 12 тысяч долларов США сегодня).
Этот металлический барвинковый синий цвет хорошо сочетается с экстерьером, искусно утонченная и красиво пропорциональная форма, написанная Альдо Бровароне под Pininfarina. Расположенный спереди двигатель h5 позволяет использовать низкий наклонный капот, который приводит к сильно наклоненному лобовому стеклу, в то время как относительно толстые стойки C обращают внимание на слегка вогнутую крышку багажника.
Состояние лакокрасочного покрытия, отделки, фар и всего остального на фотографиях хорошо сохранилось, только на бамперах видны небольшие дефекты.Маркировка полная и правильная, в то время как безупречная выхлопная труба также выглядит правильно. Колеса имеют заводские прорези 15 дюймов, обернутые в старые шины, которые будут заменены перед продажей.
Продавец описывает обивку как неоново-синий велюр, который, по нашему мнению, хорошо контрастирует с экстерьером и кремовыми дверными панелями / спинками сидений. Эти автомобили, как правило, не очень хорошо стареют внутри из-за хрупкого пластика, но этот выглядит отлично с учетом, вероятно, оригинального радио Becker и интересного рулевого колеса OEM.
Отделение багажника красиво отделано и, похоже, содержит полный набор инструментов с соответствующим заводским кожаным чехлом.